Chapter: Plant Biology : Growth and development
Molecular action of hormones and intracellular messengers
MOLECULAR ACTION OF HORMONES AND INTRACELLULAR MESSENGERS
Key Notes
How do plant hormones act?
Plant hormones act in a variety of types of responses that may be long term, such as growth and development, or rapid. Hormones alter the activity of enzymes or other cytoplasmic components directly, alter gene expression and the production of new cellular components, or both.
Receptors and target tissues
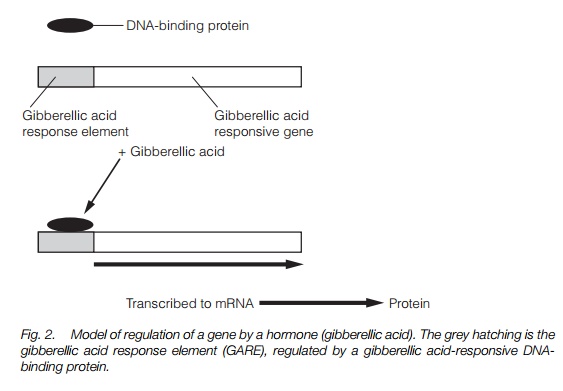
In order for a hormone response to occur, the presence of the hormone must first be perceived by a receptor protein. Receptors for ethylene (ETR1) and auxin (ABP1) have been identified and characterized. Target tissues respond to a given hormone because they possess the necessary receptors and pathways for a response.
Hormones and gene expression control
Many genes are regulated by plant hormones. Some respond very rapidly, in a matter of minutes; others require hours to days. Genes that respond to hormones have a region termed a response element in the promoter region that is regulated by a protein (transcription factor) which in turn is regulated by the hormone.
Intracellular messengers
Intracellular messengers alter in response to a stimulus causing a coordinated response within the cell. Ca2+and inositol trisphosphate (IP3) are examples. IP3 is produced when phospholipase C is activated and releases Ca2+ from intracellular stores. Intracellular messengers frequently activate protein kinases which phosphorylate other proteins
How do plant hormones act?
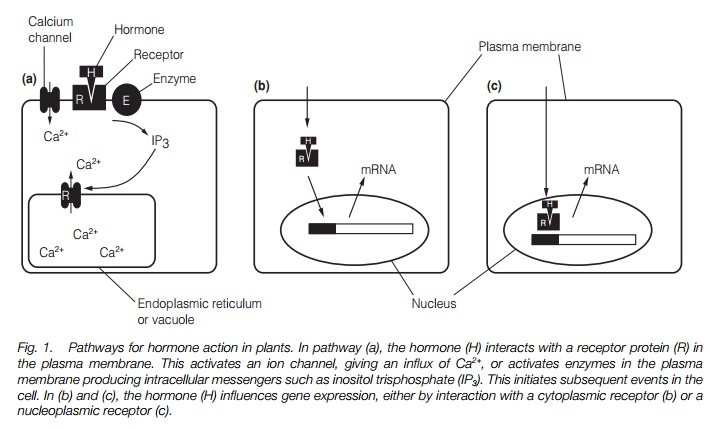
Plant hormones influence both long-term processes, such as growth and development, and short-term responses such as the closure of stomata or curvature in unilateral light or gravity. These effects may involve altered gene expression and altered activity of cellular components such as enzymes or the cytoskeleton. To do this, there must be a cellular component called a receptor that alters in function or property in response to the presence of the hormone. Between the receptor and point of action there may be an inter-linked series of chemical or ionic signals, termed intracellular messengers, within the cell that transmits and amplifies the initial signal. Plant hormones act in a variety of different ways, with the same hormone having several different effects using several different mechanisms in the same plant, and even in the same tissue. Figure 1 illustrates some of the potential pathways involved, while Table 1 details some examples of plant hormone actions and their mechanisms.
Receptors and target tissues
A receptor is a protein which binds the hormone. Binding has high affinity and specificity, but is reversible. In other words, a hormone is bound at low concentrations, but can be readily released, ‘switching’ off the response. Similar compounds not effective as hormones are not bound. Binding of the hormone produces a change in the receptor that results in it being able to activate other processes. Receptors may be located in membranes or may be in the cytoplasm.
Only a few plant hormone receptors have been identified with certainty. ABP1 is a soluble auxin binding protein that normally resides in the lumen of the endoplasmic reticulum (ER). It binds auxin reversibly, with high specificity and affinity and is a strong candidate as an auxin receptor. Anethylene receptor, ETR1, has been cloned in Arabidopsis and other plants, and has a highaffinity and specificity for ethylene. The receptor is aprotein kinase containinga copper atom as part of the ethylene-binding site. It passes on the signal byphosphorylating other proteins in a signal transduction chain (see below).While other proteins have been suggested to be receptors for hormones, nonehave yet been characterized in detail.
It is evident that some tissues respond to a hormone while others do not. Such hormone-responsive tissues are known as target tissues because they contain the receptors and signal transduction machinery necessary to respond in a particular way.
Hormones and gene expression control
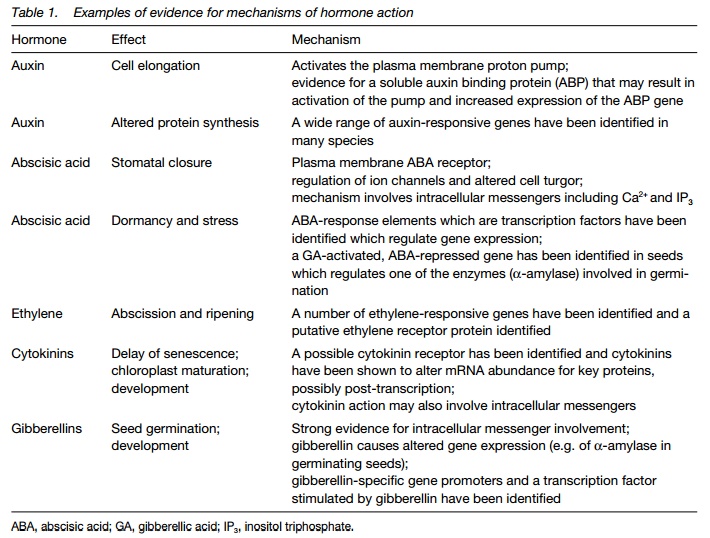
Altered gene expression is an essential component of many hormone-regulated processes. Altered gene expression results in the production of new proteins involved in the changes in cell function that hormones regulate (Fig. 2). This
may occur in addition to hormone action via intracellular messengers (see below) or by directly modulating cell processes by interaction with proteins already present. In this section, two examples will be given of gene regulation
by plant hormones: auxin and elongation growth and ethylene and fruit ripening. Hormone-responsive genes have identifiable sequences in thepromoter region (Topic B5) termed response elements that interact with protein modulators (transcription factors) which in turn are regulated by the hormone.
Auxin and elongation growth
The mechanism of the stimulation of elongation growth by auxin has been the subject of scientific controversy for many years and is still not fully understood for all plants. When oat coleoptiles are exposed to auxin, their rate of elongation increases as a result of cell elongation . The effect of auxin can be mimicked by weak acid, and it can be shown that when auxin is added to the coleoptile, the cell wall space becomes acidified. This acidification results from increased pumping of protons (H+) into the cell wall by the plasma membrane proton ATPase (proton pump; ). The acid growth theory states that the following stages occur:
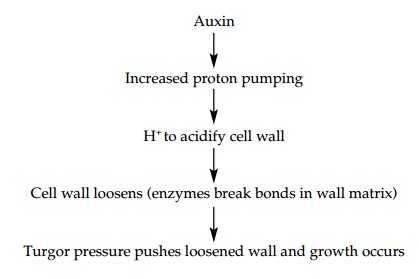
Proteins (expansins) present in the cell wall catalyze pH-dependent wall extension growth, by loosening bonds between components of the wall matrix. How does auxin increase the acidification of the cell wall? Two possibilities exist: the first is the activation of proton ATPase already present at the plasma membrane; the second is that auxin causes an increase in the amount of proton ATPase present by upregulating gene expression. In addition to the effect of auxin on the proton pump, other auxin-responsive mRNAs have been found, some of which also increase during elongation. Table 2summarizes some of these and their possible functions.’

Ethylene and fruit ripening
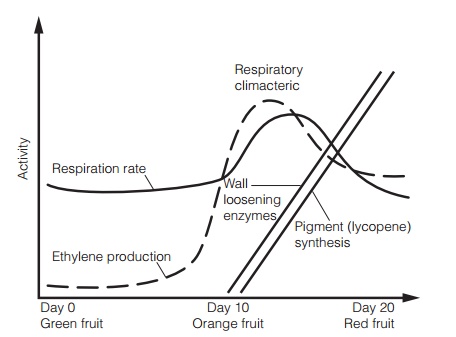
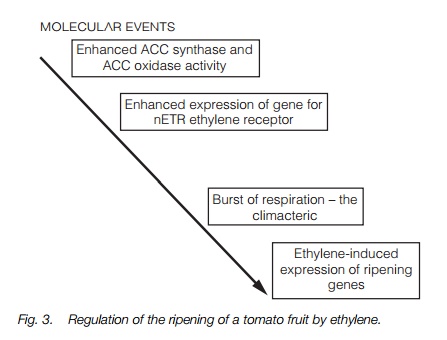
Tomato fruit ripening follows a well defined series of events typical of climacteric fruits (fruits which show a burst of respiration during ripening;Fig. 3).Ripening involves softening of the cell walls, increase in sugar content and color changes which ultimately result in an attractive red fruit. Studies of the proteins involved in ripening reveal that a well-regulated series of changes in geneexpression occurs. Ethylene synthesis increases rapidly before the climacteric as genes for the enzyme ACC oxidase (Topic F2) are expressed, and declines after it. The ethylene signal is then sensed by the ethylene receptor (ETR1) and transcription of a range of mRNAs for proteins involved in ripening is enhanced. The level of transcription of the gene for ETR1 also increases greatly during ripening. These ethylene regulated genes encode mRNAs for cell wall softening and pigment synthesis. One well studied gene, which is transcribed in response to ethylene, encodes the wall-loosening enzyme polygalacturonase (PG). Tomatoes in which expression of PG has been reduced by antisense technology show enhanced storage as they ripen more slowly than conventional tomatoes. In this technique, a single strand of DNA is present that binds specifically with the PG mRNA strand, thereby inactivating it. Such fruits also have better qualities for processing and suffer fewer losses in harvest and transport to the consumer or processing plant.
Intracellular messengers
Hormones generally convey signals from cell-to-cell and tissue-to-tissue. Intracellular messengers carry out signaling within cells. They may respond to a signal from a hormone first perceived at the plasma membrane, or they may respond to a stimulus (such as light or temperature) first perceived inside the cell. A number of intracellular signaling molecules have been found in plants; two, Ca2+ and the inositol trisphosphate (IP3) pathway will be described here.
Calcium as an intracellular messenger
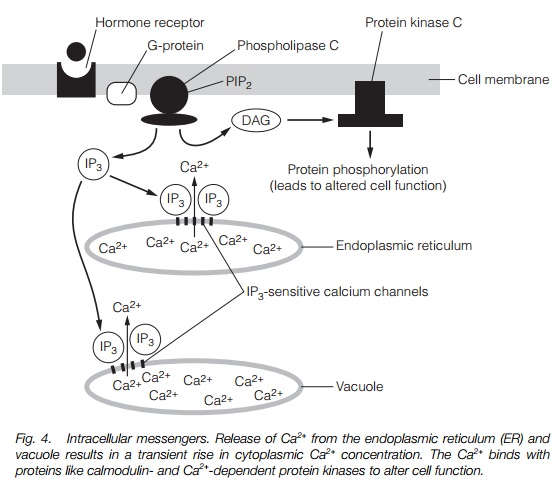
Cells maintain a very low concentration of Ca2+ in the cytoplasm (<1 M), by pumping it out across the plasma membrane to the apoplast usingCa2+ATPases. They also pump Ca2+ into intracellular stores such as the vacuole and endoplasmic reticulum. This results in a very steep Ca2+ gradient across several cell membranes. When calcium channels (Topic I3) in these membranes open, Ca2+ floods into the cytoplasm, giving a Ca2+ ‘wave’. Channel opening may be regulated by a hormone receptor or by some other stimulus. This amplifies the signal (one molecule of hormone bound can keep a channel open long enough to permit tens of Ca2+ ions to enter) and can integrate (coordinate) several signals. The Ca2+ wave is then perceived by receptor proteins. The best described of these is calmodulin, a protein with four Ca2+ binding sites. When calmodulin binds Ca2+, it changes conformation and activates a range of proteins, including Ca2+-calmodulin-dependent protein kinases (CaMPKs). Plants also have a range of Ca2+-dependent (but calmodulin independent) protein kinases. Protein kinases are important in signal transduction pathways as they phosphorylate other proteins at specific sites, altering their activity.
Inositol trisphosphate
The inositol triphosphate (IP3) pathway begins with the conversion of a plasma membrane lipid, phosphatidyl inositol into phosphatidyl inositol bisphosphate (PIP2) by kinase enzymes in the plasma membrane. PIP2 is then hydrolyzed by phospholipase C (PLC) to give IP3 and diacylglycerol(DAG). PLC activity is regulated by a signal transduction pathway initiated by the binding of a hormone to a receptor, so IP3 and DAG levels respond to stimuli.
IP3 causes Ca2+ channels to open at the tonoplast (vacuole) and endoplasmic reticulum, resulting in a Ca2+ signal being initiated (see above). DAG activates protein kinase C that regulates other processes. Figure 4 summarizes the Ca2+ and inositol trisphosphate signaling pathways.
Related Topics