Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: Oxygen, metabolism, and energetics
Metabolic rate of Fishes
Metabolic rate
Metabolism is the sum total of all biochemical processes taking place within an organism. Since these reactions give off heat as a byproduct, measuring the heat lost by an animal probably is the best way to measure its metabolism. This can be a difficult process, however, so frequently another parameter related to metabolism serves as an indirect measure. In fishes the rate of oxygen consumption is frequently used as an indicator of metabolic rate, but we must assume that no significant anaerobic metabolism takes place during the measurement period.
Metabolic rates can be influenced by a variety of factors, including age, sex, reproductive status, food in the gut, physiological stress, activity, season, and temperature. For this reason, it is useful to define metabolic terms. Standard metabolic rate is often defined as the metabolic rate of fish while it is at rest and has no food in its gut. However,Belokopytin (2004) points out that many fishes under natural conditions feed regularly and therefore almost always have some food in the gut, so some amount of digestion is likely to be part of a fish’s metabolism at all times. Fishes rarely remain still while metabolic rates are being measured, so the term routine metabolic rate is often used to indicate that the rate was measured during routine activity levels. The resulting estimates of metabolic rate are therefore higher than what might be expected for a resting fish. Sometimes researchers will measure metabolism at several levels of activity and extrapolate back to zero activity to estimate standard metabolic rate. Because metabolisms affected by temperature, the temperature should be recorded whenever measuring fish metabolism.
Metabolic rate increases with activity until a fish reaches the point at which it is using oxygen as rapidly as its uptake and delivery system can supply it. This is its maximum (or active) metabolic rate. The difference between the standard metabolic rate and the maximum metabolic rate at any given temperature is known as the metabolic scope. The concept of metabolic scope can be important in trying to understand fish’s metabolic limits. Any factors that increase standard or routine metabolic rates, such as stress due to disease, handling, reproduction, or environmental conditions, narrow this scope and may limit other activities.
In general, fishes tend to have higher metabolic rates at higher temperatures, so as temperature increases a fish’s need for oxygen also increases. Because the availability of oxygen in water decreases with increasing temperature, warm conditions stress most fishes. This stress probably was an important selection factor favoring the evolution of air breathing in many tropical fishes.
Under laboratory conditions fish acclimated to low temperatures consume less oxygen than fish of the same species acclimated to higher temperatures (see, for example, Beamish 1970; Brett 1971; Kruger & Brocksen 1978;DeSilva et al. 1986). The rates of many biochemical reactions increase with temperature, thereby increasing the need for oxygen to provide the energy needed to support increased levels of cellular metabolism. However, trends such as this observed in laboratory studies may not reflect seasonal changes in metabolic rate. Under natural environmental conditions, the gradual acclimatization of a fish to seasonal changes involves many physiological processes, each of which can have an impact on overall metabolism. Therefore, the results of temperature acclimationstudies during a single season may not represent true seasonal changes in metabolic rates (Moore & Wohlschlag 1971;Burns 1975; Evans 1984; Adams & Parsons 1998; Gamperlet al. 2002).
Temperature–metabolic rate generalizations based on studies of individual species acclimated to different temperatures should not be applied across species, especially those adapted to very different thermal environments. At low temperatures, for example, polar fishes have metabolic rates considerably higher than those of temperate species acclimated to the same low temperatures (Brett & Groves1979). Metabolic rates of tropical fishes and those of temperate species acclimated to high temperatures differ only slightly.
Size also can have a considerable effect on metabolism. Not surprisingly, large fishes generally will have higher overall metabolic rates than small fishes, assuming other factors such as activity are constant. However, the metabolic rate per unit of mass, often called the mass-specific metabolic rate or metabolic intensity, is higher for smaller fishes. This relationship seems to hold true for other animal groups as well.
Among the more metabolically costly things that a fish does is to swim. Because water is 800 times denser than air,more energy is required to move through it. There is a trade-off, however, in that the density of water also provides buoyancy so that fishes do not have to utilize as much energy fighting gravity as they would in a less dense medium. Not surprisingly, oxygen consumption in fishes increases with swimming velocity. The increase is exponential, starting out quite slowly at first, but increasing dramatically at higher velocities (Fig. 5.7). Such oxygen consumption curves probably underestimate the true metabolic cost of swimming at high speeds because of the increased use of anaerobic metabolism by swimming muscles at higher velocities.
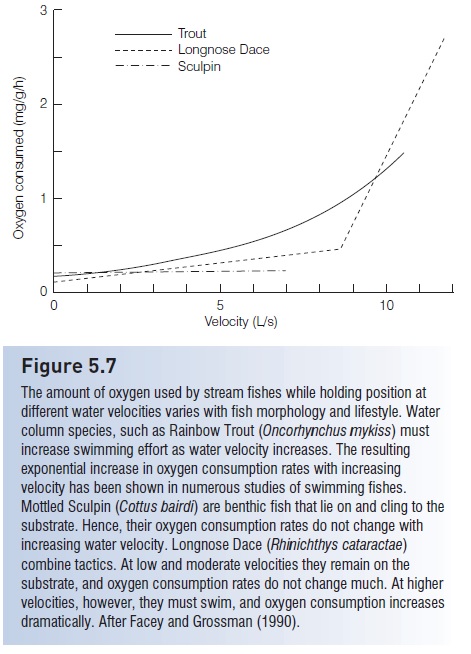
Figure 5.7
The amount of oxygen used by stream fishes while holding position at different water velocities varies with fish morphology and lifestyle. Water column species, such as Rainbow Trout (Oncorhynchus mykiss) must increase swimming effort as water velocity increases. The resulting exponential increase in oxygen consumption rates with increasing velocity has been shown in numerous studies of swimming fishes. Mottled Sculpin (Cottus bairdi) are benthic fish that lie on and cling to the substrate. Hence, their oxygen consumption rates do not change with increasing water velocity. Longnose Dace (Rhinichthys cataractae)combine tactics. At low and moderate velocities they remain on the substrate, and oxygen consumption rates do not change much. At higher velocities, however, they must swim, and oxygen consumption increases dramatically. After Facey and Grossman (1990).
The evolution of a torpedo-shaped, fusiform body undoubtedly is the result of its energetic advantages. Fin shape and placement also are important considerations, as well as body flexion during the act of swimming. The fastest, most active swimmers are streamlined, with high, thin caudal fins that oscillate rapidly while the rest of the body remains fairly rigid. This eliminates the drag that would be created by throwing most of the body into curves while swimming forward.
Body shape and other morphological features also are important to the energetics of many benthic fishes. Bottom dwelling stream fishes, for example, are able to hold their position in a high-flow environment without much energetic cost due to body shape and judicious use of their fins. Mottled Sculpin (Cottidae) can use their pelvic fins to hold to the rocky substrate of swift mountain streams. They can even hold position in a Plexiglas’s swimming tunnel, apparently by using their large pectoral fins to create downward force as the water flows over them (Facey & Grossman 1990). Their overall body shape of a large head and an arrow, tapering body may also help them remain on the bottom as water flows over them. These morphological adaptations give sculpins the ability to hold position immoderate currents without a significant energetic cost(Fig. 5.7). The bottom-foraging Longnose Dace (Cyprinidae)responds similarly at low to moderate velocities, showing no change in oxygen consumption. At higher velocities, however, it must resort to swimming to hold its position and its oxygen consumption increases dramatically. This change in behavior breaks the oxygen consumption curve into two distinct segments (Fig. 5.7).
Related Topics