Chapter: Aquaculture Principles and Practices: Reproduction and Genetic Selection
Mass selection - Genetic selection and hybridization in Aquaculture
Mass selection
Mass selection, or individual selection, is based on characteristics of
the individuals under selection as opposed to selection based on the
performance of their relatives. It is one of the simplest and most common
methods employed in breeding programmes, where the character-istic to be
improved is easy to measure. It can be used efficiently in selection for growth
rate and to some extent for age at maturity. As stated above, response in mass
selection
(R) is determined by the general equation
R = iσh2 = Sh2 where
S = selective differential (the difference in a certain trait between
the individuals selected and the population as a whole),
h2 = heritability of the differences (the share of additive genetic
variation in the general variation of the character), and
i = intensity of selection.
The high fecundity of cultivated fish causes high selective potential
and intensity of selection compared to domestic animals and poultry. In
performing mass selection in fish breeding, the selection severity coefficient
or the rejection rigidity factor (V) is calculated by the equation:
V=100n/N %
where
n =the number of individuals selected and
N =the total number of fish grown.
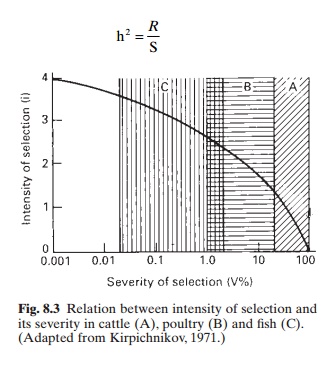
In fig. 8.3 the intensity of rejection is plotted against its severity
on a semi-logarithmic scale. The curve obtained shows that there is a sharp
increase in the intensity of selection with decrease in the severity
coefficient within the range 100–10 per cent. A further decrease in V (down to
1 per cent) results in a considerably lower increase in i; with further
decrease in V (0.1–0.01) there is hardly any increase in i. For fish with high
fecundity, selection gives best results when the severity of selection is 1–0.1
per cent.
Response to selection is directly proportional to the heritability of
the character (h2). In many cases, a rather accurate estimate of the value of
heritability of the character can be obtained from the equation:
h2 = R/S
To obtain the estimate, selection has to be conducted in several
successive generations.
For increasing response in mass selection, the values of i, s and h2 have to be increased. The value
of i can be raised in fecund fish by increasing the number of individuals and,
through this, increasing the severity of selection. Variability should relate
only to genetic and not environmental variation, as the latter is conditioned
by the non-additive genetic variation. To increase h2 values, non-related individuals
have to be crossed. Outbreeding increases the degree of heterozygosity, i.e.
increase in genetic variation, but inbreeding results in higher homozygosity. A
sufficient number of fish should be available every year for crossing, to
enable selection of fish from different crossings for breeding purposes.
Another method of increasing genetic variation is by speeding up the process of
mutation through irradiation and chemical mutagens.
Non-hereditary variation can be depressed by following special rearing
methods such as growing spawners under favourable conditions for maturation,
simultaneous crossings, incubation of all eggs under identical physico-chemical
conditions, growing larvae and young ones under conditions that do not promote
food competition, avoiding the mixing of stocks grown in different ponds or
enclosures, and by selection at the age when the animal is more susceptible to
improvement by selection.
It is necessary to point out here that a long period of one-way
selection for certain characteristics may bring about changes in other
morphogenetically or genetically correlated characters. There are many observed
examples of correlated responses in selection of non-selected characters, such
as physiological and biochemical factors (Steffens, 1964), growth rate (Moav
and Wohlfarth, 1967), fecundity, etc.
Related Topics