Chapter: Anatomy of Flowering Plants: An Introduction to Structure and Development : Leaf
Leaf Epidermis
Leaf Epidermis
The leaf epidermis is a complex tissue that usually consists of a single layer of cells, though in a few species (e.g. Ficus and Peperomia) the epidermis proliferates to form several cell layers (a multiple epidermis). At maturity it is difficult to distinguish a multiple epidermis from a hypodermis. The specialized elements of the leaf epidermis are essentially the same as those of the stem: stomata, trichomes, papillae, surface sculpturing, epicuticular wax and variously differentiated pavement epidermal cells.
1. Pavement Epidermal Cells
In surface view, pavement epidermal cells can be elongated or more or less isodiametric, and their anticlinal walls can be straight or undulating (Fig. 4.3A). Anticlinal cell walls are often more sinuous on the abaxial than the adaxial surface of the same leaf. Cells that lie over veins are often elongated in the direction of the veins. In linear leaves of the type found in many monocots, epi-dermal cells are typically elongated parallel to the long axis of the leaf (Fig. 4.3B).
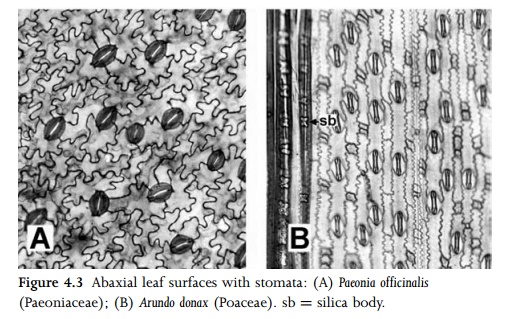
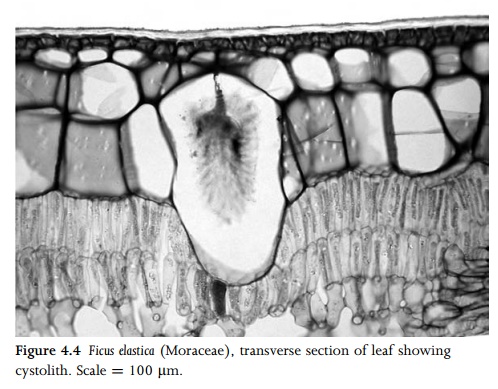
Epidermal cells frequently vary in size and wall thickness in different parts of the same leaf. In some Poaceae (e.g. Zea mays), enlarged cells (termed bulliform cells) occur in restricted regionsof the leaf epidermis (Fig. 4.9). In some species these cells play a role in the unrolling of the leaf in response to turgor pressure and water availability. Occasional epidermal cells may contain crystals or silica bodies. Cystoliths can be restricted to individual epidermal cells (e.g. in the family Opiliaceae), or they can span both the epidermis and underlying mesophyll (e.g. in Ficus: Fig. 4.4). They contain a body encrusted with calcium carbonate that is attached to the cell wall by a silicified stalk36. Silica bodies occur in the leaf epidermis of the monocot families Cyperaceae, Poaceae (Fig. 4.3B) and Arecaceae. The grass epidermis typically consists of both long and short cells, the short cells sometimes forming the bases of trichomes.
2. Stomata
Since they control gaseous exchange (water release and carbon dioxide uptake), stomatal pores occur in almost all angiosperm leaves, though their distribution varies in differ-ent species. In many species, stomata are present on both leaf surfaces (i.e. in amphistomatic leaves), whereas in others they
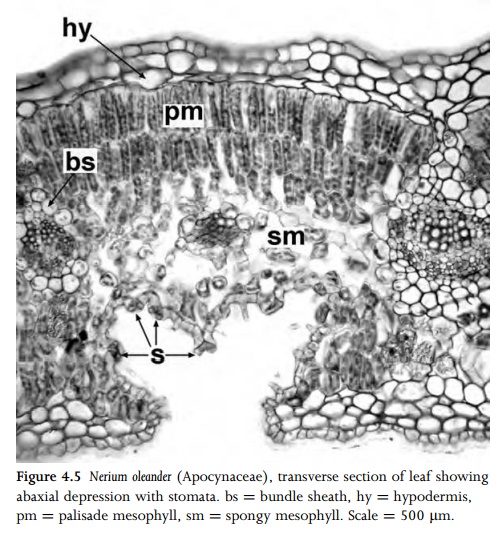
are restricted to the abaxial surface (hypostomatic leaves). Rarely, stomata are restricted to the adaxial surface (in epistomatic leaves); this condition occurs mainly on the floating leaves of aquatic plants. Within each leaf, stomata most frequently occur on regions overlying the chlorenchymatous mesophyll rather than the veins. In xeromorphic species stomata are often protected to restrict water loss; for example, individual stomata may be sunken, or groups of stomata may be restricted to hair-lined grooves or depressions on the abaxial leaf surface (Fig. 4.5). This creates a pocket of water vapour, and thus reduces water loss by transpiration.
3. Trichomes and Papillae
Similarly, the distribution of leaf-borne trichomes varies among species. Trichomes can occur on the entire leaf surface, or they can be restricted to certain areas, such as abaxial
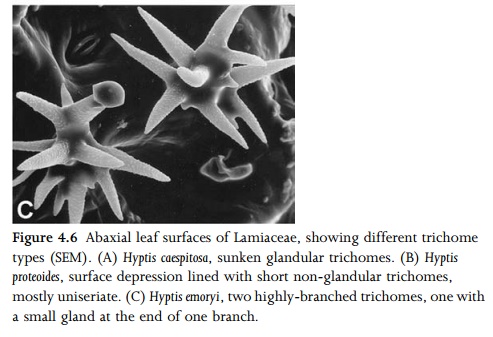
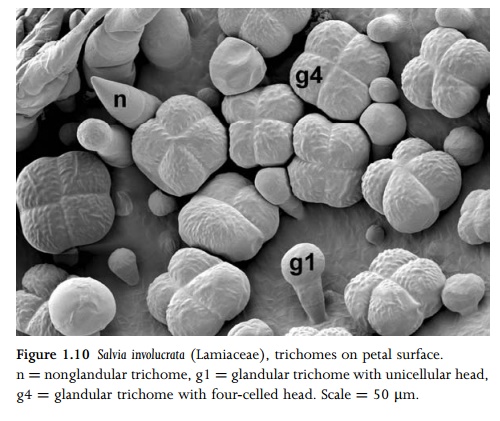
surface grooves or leaf margins. Some species possess several different types of trichome on the same leaf. For example, many species of the mint family Lamiaceae characteristically possess two or more sizes of glandular trichome and either branched or unbranched nonglandular trichomes (Figs 1.10, 4.6). Specialized hair types include stinging hairs (e.g. in Urtica dioica: Fig. 1.11), water-absorptive leaf scales (e.g. in Tillandsia), and salt-secreting glands (e.g. in Avicennia, Limonium and Tamarix). Salt glands accu-mulate sodium chloride and so allow plants to colonize highly saline soils.
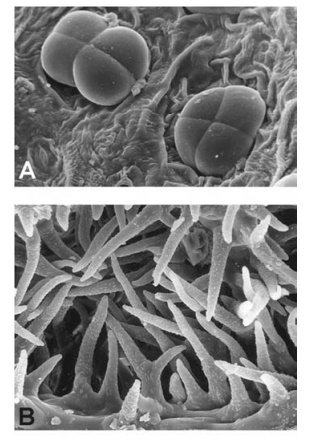
Papillae are short epidermal projections that are present on the leaf surfaces of some species. For example, within the Iris family (Iridaceae), papillae are frequently present on leaf epidermal cells, in some species as a single papilla per cell, and in other species in a row of up to nine papillae per cell. Some species possess a ring of four or more (sometimes coalesced) papillae around each stoma, together forming a raised rim.

4. Cuticle and Wax
The cuticle is a non-cellular layer composed of several inert poly-mers, especially cutin, that covers the entire leaf surface and most other aerial plant surfaces. In mesomorphic leaves the cuticle is typically thin and almost transparent, but many xeromorphic plants possess a thick leaf cuticle that often appears lamellated in transverse section. In some species the outer surface of the cuticle possesses characteristic patterns of ridges, folds or striations. These can be short or long and oriented randomly or in a regular pattern, sometimes radiating around stomata or trichomes. Surface pat-terning may have biological significance in relation to mechanical and optical properties and wettability of the surface. For example, the cuticular patterns on the mature leaf surfaces of Aloe and related genera are under precise genetic control.
Leaves of some species have a covering of wax over the cuticle. Epicuticular wax is either in the form of a surface crust, or more commonly in small particles of varying shapes and sizes, ranging from flakes to filaments and granules. Wax particles are vari-ously orientated and sometimes occur in characteristic patterns. Certain compounds, such as terpenes and flavonoids, can be recognized by their detailed structure.
Related Topics