Chapter: Medical Surgical Nursing: Assessment of Immune Function
Immune Function: Defenses and Responses
IMMUNE
FUNCTION: DEFENSES AND RESPONSES
There are two general types of immunity:
natural (innate) and ac-quired (adaptive). Natural immunity is a nonspecific
immunity present at birth. Acquired or specific immunity develops after birth.
Natural immune responses to a foreign invader are very similar from one
encounter to the next regardless of the number of times the invader is
encountered; in contrast, acquired re-sponses increase in intensity with
repeated exposure to the in-vading agent (Delves & Roitt, 2000a). Although
each type of immunity plays a distinct role in defending the body against
harmful invaders, the various components usually act in an inter-dependent
manner.
Natural Immunity
Natural (innate) immunity provides a
nonspecific response to any foreign invader, regardless of the invader’s
composition. The basis of natural defense mechanisms is the ability to
distinguish between friend and foe or “self” and “nonself.” Such natural
mechanisms include
physical and chemical barriers, the action of WBCs, and inflammatory responses.
PHYSICAL AND CHEMICAL BARRIERS
Physical surface barriers include intact skin
and mucous mem-branes, which prevent pathogens from gaining access to the body,
and the cilia of the respiratory tract along with coughing and sneezing
responses, which act to filter and clear pathogens from the upper respiratory
tract before they can invade the body fur-ther. Chemical barriers, such as
acidic gastric secretions, mucus, enzymes in tears and saliva, and substances
in sebaceous and sweat secretions, act in a nonspecific way to destroy invading
bac-teria and fungi. Viruses are countered by other means, such as in-terferon.
Interferon, one type of biologic
response modifier, is a nonspecific viricidal protein naturally produced by the
body that is capable of activating other components of the immune system.
WHITE BLOOD CELL ACTION
WBCs,
or leukocytes, participate in both the natural and the ac-quired immune
responses. Granular leukocytes, or granulocytes (so called because of granules
in their cytoplasm), fight invasion by foreign bodies or toxins by releasing
cell mediators, such as his-tamine, bradykinin, and prostaglandins, and
engulfing the foreign bodies or toxins. Granulocytes include neutrophils,
eosinophils, and basophils.
Neutrophils (also called polymorphonuclear
leukocytes, or PMNs, because their nuclei have multiple lobes) are the first
cells to arrive at the site where inflammation occurs. Eosinophils and
ba-sophils, other types of granulocytes, increase in number during allergic
reactions and stress responses. Nongranular leukocytes in-clude monocytes or
macrophages (referred to as histiocytes when they enter tissue spaces) and
lymphocytes. Monocytes also function as phagocytic
cells, engulfing, ingesting, and destroying greater numbers and quantities
of foreign bodies or toxins than granulo-cytes. Lymphocytes, consisting of B
cells and T cells, play major roles in humoral and cell-mediated immune
responses. About 60% to 70% of lymphocytes in the blood are T cells, and about
10% to 20% are B cells (Porth, 2002).
INFLAMMATORY RESPONSE
The
inflammatory response is a major function of the natural (non-specific or
innate) immune system elicited in response to tissue in-jury or invading
organisms. Chemical mediators assist this response by minimizing blood loss,
walling off the invading organism, acti-vating phagocytes, and promoting
formation of fibrous scar tissue and regeneration of injured tissue.
Dysfunction
of the natural immune system can occur when the immune components are
inactivated or when they remain active long after their effects are beneficial.
Immunodeficiencies are char-acterized by inactivation or impairment of immune
components, and disorders with an inflammatory component (eg, asthma, allergy,
arthritis) are characterized by persistent inflammatory responses (Medahitov
& Janeway, 2000). The immune system’s recognition of one’s own tissues as
“foreign” rather than as self is the basis for many autoimmune disorders.
Acquired Immunity
Acquired (adaptive) immunity—immunologic responses acquired during life but not present at birth—usually develops as a result of prior exposure to an antigen through immunization (vaccination) or by contracting a disease, both of which generate a protective immune response. Weeks or months after exposure to the disease or vaccine, the body produces an immune response that is sufficient to defend against the disease upon re-exposure to it.
The two types of acquired immunity are known
as active and passive. In active acquired immunity, the immunologic defenses
are developed by the person’s own body. This immunity gener-ally lasts many
years or even a lifetime.
Passive acquired immunity is temporary
immunity transmit-ted from another source that has developed immunity through
previous disease or immunization. For example, immune globu-lin and antiserum,
obtained from the blood plasma of people with acquired immunity, are used in
emergencies to provide immu-nity to diseases when the risk for contracting a specific
disease is great and there is not enough time for a person to develop ade-quate
active immunity. For example, immune globulin may be administered to those
exposed to hepatitis. Immunity resulting from the transfer of antibodies from
the mother to an infant in utero or through breastfeeding is another example of
passive im-munity. Active and passive acquired immunity involve humoral and
cellular (cell-mediated) immunologic responses (Ada, 2001).
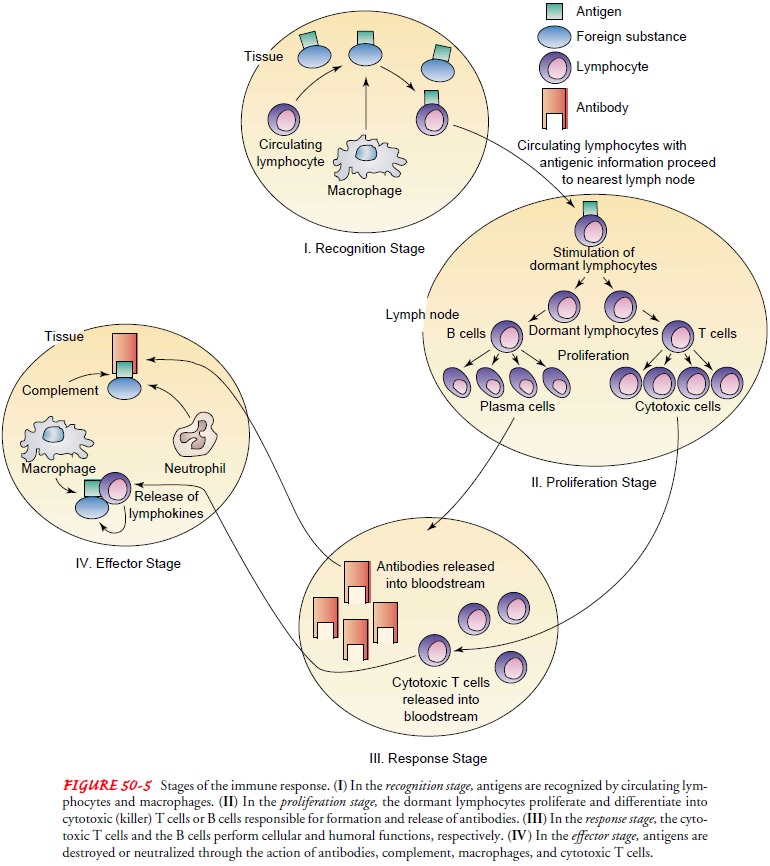
Response to Invasion
When
the body is invaded or attacked by bacteria, viruses, or other pathogens, it
has three means of defending itself:
· The phagocytic immune
response
· The humoral or antibody
immune response
· The cellular immune
response
The first line of defense, the phagocytic immune response, involves
the WBCs (granulocytes and macrophages), which have the ability to ingest
foreign particles. These cells move to the point of attack, where they engulf
and destroy the invading agents. Phagocytes also remove the body’s own dying or
dead cells. Cells in necrotic tissue that are dying release substances that
trigger an inflammatory response. Apoptosis,
or programmed cell death, is the body’s way of destroying unwanted cells such
as cancer cells or cells that die a natural death. Apoptosis involves the
digestion of DNA by endonucleases, resulting in the cells being targeted for
phagocytosis (Delves & Roitt, 2000a).
Unlike
macrophages, eosinophils are only weakly phagocytic. On activation, eosinophils
probably kill parasites by releasing spe-cific chemical mediators into the
extracellular fluid. Additionally, they secrete leukotrienes, prostaglandins,
and various cytokines (Delves & Roitt, 2000a).
A second protective response, the humoral immune response (sometimes called the antibody response), begins with the B lym-phocytes, which can transform themselves into plasma cells that manufacture antibodies. These antibodies, highly specific pro-teins, are transported in the bloodstream and attempt to disable the invaders. The third mechanism of defense, the cellular im-mune response, also involves the T lymphocytes, which can turninto special cytotoxic (or killer) T cells that can attack the patho-gens themselves.
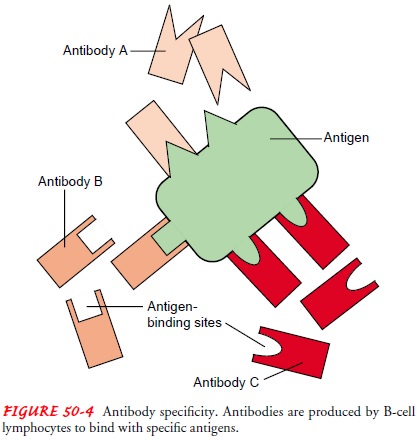
The
part of the invading or attacking organism that is respon-sible for stimulating
antibody production is called an antigen
(or an immunogen). For example, an antigen can be a small patch of proteins on
the outer surface of the microorganism. Not all anti-gens are naturally
immunogenic and must be coupled to other molecules to stimulate the immune
response. A single bacterium, even a single large molecule, such as a toxin
(diphtheria or teta-nus toxin), may have several such antigens, or markers, on
its sur-face, thus inducing the body to produce a number of different
antibodies. Once produced, an antibody is released into the bloodstream and
carried to the attacking organism. There it com-bines with the antigen, binding
with it like an interlocking piece of a jigsaw puzzle (Fig. 50-4). There are
four well-defined stages in an immune response: recognition, proliferation,
response, and effector.
RECOGNITION STAGE
Recognition of antigens as foreign, or nonself, by the immune system is the initiating event in any immune response. The body must first recognize invaders as foreign before it can react to them.
The body
accomplishes recognition using lymph nodes and lymphocytes for surveillance.
Lymph nodes are widely dis-tributed internally throughout the body and in the
circulating blood, and externally near the body’s surfaces. They continuously
discharge small lymphocytes into the bloodstream. These lym-phocytes patrol the
tissues and vessels that drain the areas served by that node.
Lymphocytes
recirculate from the blood to lymph nodes and from the lymph nodes back into
the bloodstream, in a never-ending series of patrols. Some circulating
lymphocytes can sur-vive for decades. Some of these small, hardy cells maintain
their solitary circuits for the person’s lifetime.
The exact way in which circulating lymphocytes recognize antigens on foreign surfaces is not known; however, recogni-tion is thought to depend on specific receptor sites on the sur-face of the lymphocytes. Macrophages play an important role in helping the circulating lymphocytes process the antigens.
Both macrophages and
neutrophils have receptors for antibodies and complement; as a result, the
coating of microorganisms with antibodies, complement, or both enhances
phagocytosis. The engulfed microorganisms are then subjected to a wide range of
toxic intracellular molecules. When foreign materials enter the body, a
circulating lymphocyte comes into physical contact with the surfaces of these
materials. Upon contact, the lym-phocyte, with the help of macrophages, either
removes the anti-gen from the surface or in some way picks up an imprint of its
structure, which comes into play with subsequent re-exposure to the antigen.
In a streptococcal throat infection, for
example, the strepto-coccal organism gains access to the mucous membranes of
the throat. A circulating lymphocyte moving through the tissues of the neck
comes in contact with the organism. The lymphocyte, familiar with the surface
markers on the cells of its own body, rec-ognizes the antigens on the microbe
as different (nonself) and the streptococcal organism as antigenic (foreign).
This triggers the second stage of the immune response—proliferation.
PROLIFERATION STAGE
The circulating lymphocyte containing the
antigenic message re-turns to the nearest lymph node. Once in the node, the
sensitized lymphocyte stimulates some of the resident dormant T and B
lym-phocytes to enlarge, divide, and proliferate. T lymphocytes dif-ferentiate
into cytotoxic (or killer) T cells, whereas B lymphocytes produce and release
antibodies. Enlargement of the lymph nodes in the neck in conjunction with a
sore throat is one example of the immune response.
RESPONSE STAGE
In the
response stage, the changed lymphocytes function either in a humoral or a
cellular fashion. The production of antibodies by the B lymphocytes in response
to a specific antigen begins the humoral response. Humoral refers to the fact
that the antibodies are released into the bloodstream and so reside in the
plasma (fluid fraction of the blood).
With the initial cellular response, the
returning sensitized lym-phocytes migrate to areas of the lymph node (other
than those areas containing lymphocytes programmed to become plasma cells).
Here, they stimulate the residing lymphocytes to become cells that will attack
microbes directly rather than through the ac-tion of antibodies. These
transformed lymphocytes are known as cytotoxic (killer) T cells. The T stands
for thymus, signifying that during embryologic development of the immune
system, these T lymphocytes spent time in the thymus of the developing fetus,
where they were genetically programmed to become T lympho-cytes rather than the
antibody-producing B lymphocytes. Viral rather than bacterial antigens induce a
cellular response. This re-sponse is manifested by the increasing number of T
lymphocytes (lymphocytosis) seen in the blood smears of people with viral
ill-nesses, such as infectious mononucleosis.
Most immune responses to antigens involve
both humoral and cellular responses, although one usually predominates. For
exam-ple, during transplantation rejection, the cellular response
pre-dominates, whereas in the bacterial pneumonias and sepsis, the humoral
response plays the dominant protective role (Chart 50-1).
EFFECTOR STAGE
In the effector stage, either the antibody of
the humoral response or the cytotoxic (killer) T cell of the cellular response
reaches and couples with the antigen on the surface of the foreign invader. The
coupling initiates a series of events that in most instances re-sults in the
total destruction of the invading microbes or the com-plete neutralization of
the toxin. The events involve an interplay of antibodies (humoral immunity),
complement, and action by the cytotoxic T cells (cellular immunity). Figure
50-5 summarizes the stages of the immune response.
Humoral Immune Response
The
humoral response is characterized by production of anti-bodies by the B
lymphocytes in response to a specific antigen. Although the B lymphocyte is
ultimately responsible for the pro-duction of antibodies, both the macrophages
of natural immu-nity and the special T-cell lymphocytes of cellular immunity
are involved in recognizing the foreign substance and in producing antibodies.
ANTIGEN RECOGNITION
Several theories exist about the mechanisms
by which the B lym-phocytes recognize the invading antigen and respond by
produc-ing antibodies. This is probably because the B lymphocytes recognize
invading antigens in more than one way and respond in several ways as well.
Additionally, the B lymphocytes appear to respond to some antigens by
triggering antibody formation di-rectly. In response to other antigens,
however, they need the assis-tance of T cells to trigger antibody formation.
T
cells (or T lymphocytes), part of a surveillance system dis-persed throughout
the body, recycle through the general circu-lation, tissues, and lymphatic
system. With the assistance of macrophages, the T lymphocytes are believed to
recognize the antigen of a foreign invader. The T lymphocyte picks up the
anti-genic message, or “blueprint,” of the antigen and returns to the nearest
lymph node with that message.
Production of B Lymphocytes.
B lymphocytes stored in thelymph nodes are subdivided into thousands of
clones, each re-sponsive to a single group of antigens having almost identical
characteristics. When the antigenic message is carried back to the lymph node,
specific clones of the B lymphocyte are stimulated to enlarge, divide,
proliferate, and differentiate into plasma cells capable of producing specific
antibodies to the antigen. Other lymphocytes differentiate into B-lymphocyte
clones with a memory for the antigen. These memory cells are responsible for
the more exaggerated and rapid immune response in a person who is repeatedly
exposed to the same antigen.
ROLE OF ANTIBODIES
Antibodies are large proteins called
immunoglobulins because they are found in the globulin fraction of the plasma
proteins. All im-munoglobulins are glycoproteins and contain a certain amount
of carbohydrate. The carbohydrate concentration, which ranges from
approximately 3% to 13%, is dependent upon the class of the anti-body. Each
antibody molecule consists of two subunits, each of which contains a light and
a heavy peptide chain (Fig. 50-6). The subunits are held together by a chemical
link composed of disul-fide bonds. Each subunit has a portion that serves as a
binding site for a specific antigen referred to as the Fab fragment. This site
pro-vides the “lock” portion that is highly specific for an antigen. An
additional portion, known as the Fc fragment, allows the antibody molecule to
take part in the complement system.
Antibodies defend against foreign invaders in several ways, and the type of defense employed depends on the structure and composition of both the antigen and the immunoglobulin. The antibody molecule has at least two combining sites, or Fab frag-ments. One antibody can act as a cross-link between two anti-gens, causing them to bind or clump together. This clumping effect, referred to as agglutination, helps clear the body of the in vading organism by facilitating phagocytosis. Some antibodies as-sist in removing offending organisms through opsonization. In this process, the antigen–antibody molecule is coated with a sticky substance that also facilitates phagocytosis.
Antibodies
also promote the release of vasoactive substances, such as histamine and
slow-reacting substance, two of the chem-ical mediators of the inflammatory
response.
Antibodies
do not function in isolation but rather mobilize other components of the immune
system to defend against the invader. Their usual role is to focus components
of the natural immune system on the invader. This includes activation of the
complement system and activation of phagocytosis (Delves & Roitt, 2000a).
Types of Immunoglobulins.
The body can
produce five differ-ent types of immunoglobulins. (Immunoglobulins are
com-monly designated by the abbreviation Ig.) Each of the five types, or
classes, is identified by a specific letter of the alphabet (IgA, IgD, IgE,
IgG, and IgM). Classification is based on the chemi-cal structure and biologic
role of the individual immunoglobu-lin. The following list summarizes major
characteristics of the immunoglobulins:
IgG
(75% of Total Immunoglobulin)
· Appears in serum and tissues
(interstitial fluid)
· Assumes a major role in
bloodborne and tissue infections
· Activates the complement
system
· Enhances phagocytosis
· Crosses the placenta
IgA
(15% of Total Immunoglobulin)
· Appears in body fluids
(blood, saliva, tears, breast milk, and pulmonary, gastrointestinal, prostatic,
and vaginal secretions)
· Protects against
respiratory, gastrointestinal, and genitouri-nary infections
· Prevents absorption of
antigens from food
· Passes to neonate in
breast milk for protection
IgM
(10% of Total Immunoglobulin)
· Appears mostly in
intravascular serum
· Appears as the first
immunoglobulin produced in response to bacterial and viral infections
· Activates the complement
system
IgD
(0.2% of Total Immunoglobulin)
· Appears in small amounts
in serum
· Possibly influences
B-lymphocyte differentiation, but role is unclear
IgE
(0.004% of Total Immunoglobulin)
· Appears in serum
· Takes part in allergic
and some hypersensitivity reactions
· Combats parasitic
infections
ANTIGEN–ANTIBODY BINDING
The portion of the antigen involved in binding with the antibody is referred to as the antigenic determinant. The binding of the Fab fragment (antibody-binding site) to the antigenic determi-nant can be likened to a lock-and-key situation (Fig. 50-7). The most efficient immunologic responses occur when the antibody and antigen fit exactly. Poor fit can occur with an antibody that was produced in response to a different antigen. This phenome-non is known as cross-reactivity. For example, in acute rheumatic fever, the antibody produced against Streptococcus pyogenes in the upper respiratory tract may cross-react with the patient’s heart tis-sue, leading to heart valve damage.
Cellular Immune Response
Whereas
the B lymphocytes are responsible for humoral immu-nity, the T lymphocytes (or
T cells) are primarily responsible for cellular immunity. Stem cells
continuously migrate from the bone marrow to the thymus gland, where they
develop into T cells. T cells continue to develop in the thymus gland, despite
partial degen-eration of the thymus gland that occurs at puberty (Delves &
Roitt, 2000a). By spending time in the thymus, these cells are programmed to
become T cells rather than antibody-producing B lymphocytes. Several types of T
cells exist, each with designated roles in the defense against bacteria,
viruses, fungi, parasites, and malignant cells. T cells attack foreign invaders
directly rather than by producing antibodies.
Cellular
reactions are initiated by the binding of an antigen with an antigen receptor
located on the surface of a T cell. This may occur with or without the
assistance of macrophages. The T cells then carry the antigenic message, or
blueprint, to the lymph nodes, where the production of other T cells is
stimulated. Some T cells remain in the lymph nodes and retain a memory for the
antigen. Other T cells migrate from the lymph nodes into the general
circulatory system and ultimately to the tissues, where they remain until they
either come in contact with their respec-tive antigens or die.
ROLE OF T LYMPHOCYTES
Two
major categories of effector T cells are helper T cells and cytotoxic T cells.
These cells participate in destroying foreign organisms. Other T cells include
suppressor T cells and memory T cells. T cells interact closely with B cells,
indicating that hu-moral and cellular immune responses are not separate,
unrelated processes but rather branches of the immune response that can and do
affect each other.
Helper T cells are activated upon recognition of antigens andstimulate the rest of the
immune system. When activated, helper T cells secrete cytokines that attract and activate B cells, cytotoxic T cells,
natural killer cells, macrophages, and other cells of the im-mune system.
Separate subpopulations of helper T cells produce different types of cytokines
and determine whether the immune re-sponse will be the production of antibodies
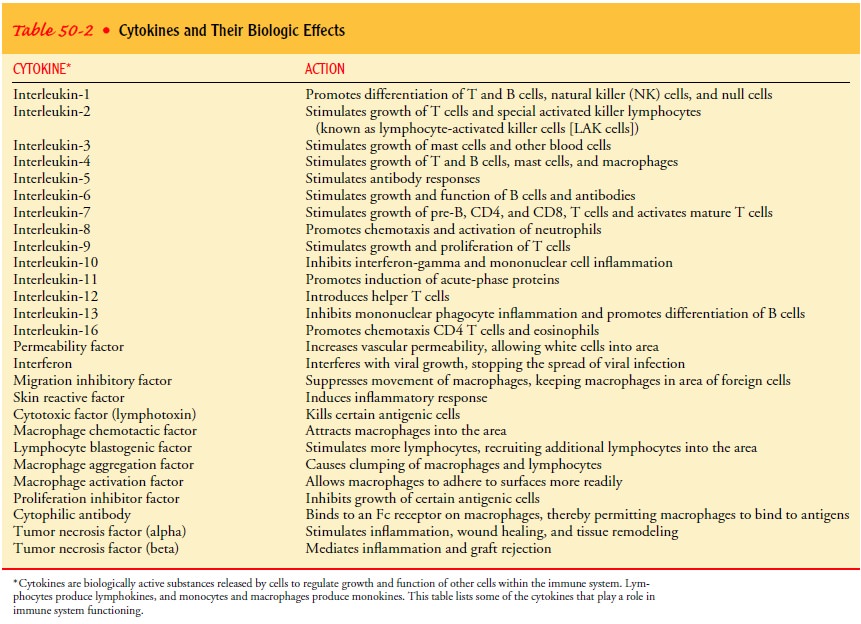
or a cell-mediated immune response. Helper T cells produce lymphokines, one category of cytokines. These lymphokines activate
other T cells (interleukin-2 [IL-2]), natural and cytotoxic T cells
(interferon-gamma), and other inflammatory cells (tumor necrosis factor).
Helper T cells produce IL-4 and IL-5, lymphokines that activate B cells to grow
and differentiate (Table 50-2).
Cytotoxic T cells (killer T cells) attack the antigen directly byaltering the cell
membrane and causing cell lysis (disintegration) and releasing cytolytic
enzymes and cytokines. Lymphokines can recruit, activate, and regulate other
lymphocytes and WBCs. These cells then assist in destroying the invading
organism. Delayed-type hypersensitivity is an example of an immune reaction
that protects the body from antigens through the production and re-lease of
lymphokines and is discussed in more detail later.
Another type of cell, the suppressor T cell, has the ability to
decrease B-cell production, thereby keeping the immune response at a level that
is compatible with health (eg, sufficient to fight in-fection adequately
without attacking the body’s healthy tissues). Memory cells are responsible for recognizing antigens from
pre-vious exposure and mounting an immune response (Table 50-3).

ROLES OF NULL LYMPHOCYTES AND NATURAL KILLER CELLS
Null
lymphocytes and natural killer (NK) cells are other lympho-cytes that assist in
combating organisms. These are distinct from B cells and T cells and lack the
usual characteristics of B cells and T cells. Null lymphocytes, a subpopulation of lymphocytes, de-stroy antigens
already coated with antibody. These cells have spe-cial Fc receptor sites on
their surfaces that allow them to couple with the Fc end of antibodies
(antibody-dependent, cell-mediated cytotoxicity).
Natural killer cells, another subpopulation of lymphocytes,defend
against microorganisms and some types of malignant cells. NK cells are capable
of directly killing invading organisms and producing cytokines. The helper T
cells contribute to the differ-entiation of null and NK cells.
Complement System
Circulating
plasma proteins, which are made in the liver and ac-tivated when an antibody
couples with its antigen, are known as complement.
These proteins interact sequentially with one an-other in a cascade or “falling
domino” effect. This complement cascade alters the cell membranes on which
antigen and antibody complex forms, permitting fluid to enter the cell and
leading eventually to cell lysis and death. In addition, activated comple-ment
molecules attract macrophages and granulocytes to areas of antigen–antibody
reactions. These cells continue the body’s de-fense by devouring the antibody-coated
microbes and by releas-ing bacterial agents.
Complement plays an important role in the immune response. Destruction of an invading or attacking organism or toxin is not achieved merely by the binding of the antibody and antigens; it also requires activation of complement, the arrival of killer T cells, or the attraction of macrophages. Complement has three major phys-iologic functions: defending the body against bacterial infection, bridging natural and acquired immunity, and disposing of im-mune complexes and the byproducts associated with inflammation (Walport, 2001a). Complement-mediated immune responses are summarized in Table 50-4.
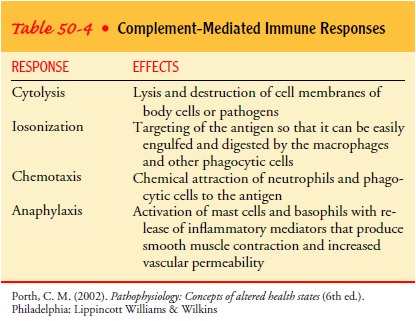
There
are several ways to activate the complement system: the classic pathway, the
alternate pathway, and the lectin pathway (Delves & Roitt, 2000a).
CLASSIC PATHWAY OF COMPLEMENT ACTIVATION
The
classic pathway (the first method discovered) is activated by antigen–antibody
complexes; it begins when antibody binds to a cell surface and ends with lysis
of the cell. It involves the reaction of the first of the circulating
complement proteins (C1) with the receptor site of the Fc portion of an
antibody molecule after for-mation of an antigen–antibody complex. The
activation of the first complement component then activates all the other
compo-nents in the following sequence: C4, C2, C3, C5, C6, C7, C8, and C9. (The components are named in the sequence
in which they were discovered.)
ALTERNATE AND LECTIN PATHWAYS
The alternate and lectin pathways of
complement activation are activated without the formation of antigen–antibody
complexes. These pathways can be initiated by the release of bacterial
prod-ucts, such as endotoxins. When complement is activated with-out the
formation of antigen–antibody complexes, the process bypasses the first three
components (C1,
C4,
and C2)
and begins with C3. Whatever the method of activation, once activated, the complement
destroys cells by altering or damaging the cell mem-brane of the antigen, by
chemically attracting phagocytes to the antigen (chemotaxis), and by rendering
the antigen more vul-nerable to phagocytosis (opsonization). The complement
sys-tem enhances the inflammatory response by releasing vasoactive substances.
Complement
components, prostaglandins, leukotrienes, and other inflammatory mediators all
contribute to the recruitment of inflammatory cells, as do chemokines, a group
of cytokines. The activated neutrophils pass through the vessel walls to
accumulate at the site of infection, where they phagocytose complement-coated
microbes (Delves & Roitt, 2000a).
This response is usually therapeutic and can be lifesaving if the cell attacked by the complement system is a true foreign invader, such as a streptococcal or staphylococcal organism. If that cell, however, is in reality part of the person—a cell of the brain or liver, the tissue lining the blood vessels, or the cells of a trans-planted organ or skin graft, for example—the result can be dev-astating disease and even death. The result of the immune response—the vigorous attack on any material identified as for-eign, the deadliness of the struggle—is obvious in the purulent material, or pus (the remains of microbes, granulocytes, macro-phages, T-cell lymphocytes, plasma proteins, complement, and antibodies), that accumulates in wound infections and abscesses. In addition, many autoimmune diseases (ie, systemic lupus ery-thematosus) and disorders characterized by chronic infection (ie, hepatitis C, bacterial endocarditis) and necrosis (myocardial infarction, stroke) are thought to be due in part to continued or chronic activation of complement, which in turn causes chronic inflammation (Walport, 2001b).
The red blood cells (erythrocytes) and platelets (thrombo-cytes) also have a role in the immune response. Red blood cells and platelets have complement receptors and as a result play an important role in the clearance of immune complexes that con-sist of antigen, antibody, and components of the complement sys-tem (Delves & Roitt, 2000a).
Role of Interferons
Biologic response modifiers, such as the
interferons, are under investigation to determine their roles in the immune
system and their potential therapeutic effects in disorders characterized by
dis-turbed immune responses. Interferons have antiviral and antitu-mor
properties. In addition to responding to viral infection, they are produced by
T lymphocytes, B lymphocytes, and macrophages in response to antigens. They are
thought to modify the immune response by suppressing antibody production and
cellular immu-nity. They also facilitate the cytolytic role of macrophages and
NK cells. Interferons are undergoing extensive testing to evaluate their
effectiveness in treating tumors and acquired immunodeficiency syndrome (AIDS).
Some interferons are already used to treat immune-related disorders (eg,
multiple sclerosis) and chronic inflammatory conditions (eg, chronic
hepatitis).
Related Topics