Chapter: Medical Microbiology: An Introduction to Infectious Diseases: Characteristics of Fungi
General Nature of Fungi
GENERAL NATURE OF FUNGI
Fungi are eukaryotes with a higher level of biological complexity than bacteria. They may be unicellular or may differentiate and become multicellular by the development of branching filaments. They reproduce sexually or asexually. The mycoses vary greatly in their manifestations but tend to be subacute to chronic with indolent, relapsing features. Acute disease, such as that produced by many viruses and bacteria, is uncommon with fungal infections.
STRUCTURE
The fungal cell has typical eukaryotic features, including a nucleus with a nucleolus, nu-clear membrane, and linear chromosomes. The cytoplasm contains an actin cytoskeleton and organelles, such as mitochondria and the Golgi apparatus. Fungal cells, which have a rigid cell wall external to the cytoplasmic membrane, differ from mammalian cells. The composition of that wall makes fungi different from bacteria and plants. Another impor-tant difference from mammalian cells involves the sterol makeup of the cytoplasmic membrane. In fungi, the dominant sterol is ergosterol; in mammalian cells, it is choles-terol. Fungi are usually in the haploid state, although diploid nuclei are formed through nuclear fusion in the process of sexual reproduction.
The chemical structure of the cell wall in fungi is markedly different from that of bacterial cells in that it does not contain peptidoglycan, glycerol or ribitol teichoic acids, or lipopolysaccharide. In their place are the polysaccharides mannan, glucan, and chitin in close association with each other and with structural proteins (Fig 45–1).
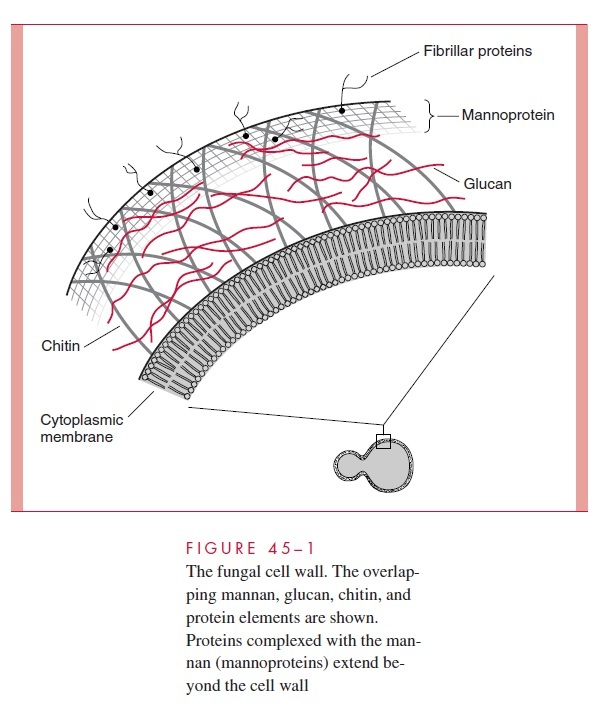
Mannoproteins are mannose-based polymers (mannan) found on the surface and in the structural matrix of the cell wall, where they are linked to protein. They are major deter-minants of serologic specificity because of variations in the composition and linkages of the polymer side chains. Glucans are glucosyl polymers, some of which form fibrils that increase the strength of the fungal cell wall, often in close association with chitin. Chitin is composed of long, unbranched chains of poly-N-acetylglucosamine. It is inert, insoluble, and rigid and provides structural support in a manner analogous to the chitin in crab shells or cellulose in plants. It is a major component of the cell wall of filamen-tous fungi. In yeasts, chitin appears to be of most importance in forming cross-septa and the channels through which nuclei pass from mother to daughter cells during cell division.
METABOLISM
Fungal metabolism is heterotrophic, requiring exogenous carbon for growth. Metabolic diversity is great, but most fungi grow with only an organic carbon source and ammo-nium or nitrate ions as a nitrogen source. In nature, nutrients for free-living fungi are de-rived from decaying organic matter. A major difference between fungi and plants is that fungi lack photosynthetic energy-producing mechanisms. Most are strict aerobes, al-though some can grow under anaerobic conditions. None are strict anaerobes.
REPRODUCTION
Fungi may reproduce by either asexual or sexual processes. Reproductive elements pro-duced asexually are termed conidia. Those produced sexually are termed spores(e.g., as-cospores, zygospores, basidiospores). Asexual reproduction involves mitotic division of the haploid nucleus and is associated with production by budding spore-like conidia or separation of hyphal elements. In sexual reproduction, the haploid nuclei of donor and re-cipient cells fuse to form a diploid nucleus, which may then divide by classical meiosis. Some of the four resulting haploid nuclei may be genetic recombinants and may undergo further division by mitosis. Highly complex specialized structures may be involved. De-tailed study of this process in fungal species such as Neurospora crassa has been impor-tant in gaining an understanding of basic cellular genetic mechanisms.
Related Topics