Chapter: Human Neuroanatomy(Fundamental and Clinical): Further Consideration of the Cerebral Cortex
Further Consideration of Functional Areas of the Cerebral Cortex
Further Consideration of Functional Areas of the Cerebral Cortex
The classical concept of the functional areas of the cerebral cortex has been presented on before. Recent work has shown that these concepts need modification in some areas and elaboration in others.
According to classical teaching the cerebral cortex has been divided into the following:
a) Motor areas
a. When these areas are stimulated electrically movements occur in various parts of the body. Anatomically, these areas give origin to projection fibres that form the corticospinal and corticonuclear tracts.
b) Sensory areas
a. Electrical activity can be recorded in these areas of cortex when a suitable sensory stimulus is applied to a part of the body. Stimulation of these areas of cortex can produce certain sensations in corresponding parts of the body. These sensory areas of the cortex receive fibres from thalamic nuclei in which major sensory pathways terminate.
c) Association areas
Areas in which a direct motor or sensory function cannot be demonstrated have been considered to be association areas where information from various sources is presumably integrated, and used to direct appropriate responses.
More recent investigations have shown that the division of the cortex into motor, sensory and association areas is an over-simplification. Many areas previously described as association areas are now known to have ‘motor’ or ‘sensory’ functions. Further it has been shown that motor and sensory functions overlap in the same areas of the cortex.
Motor and Premotor Areas
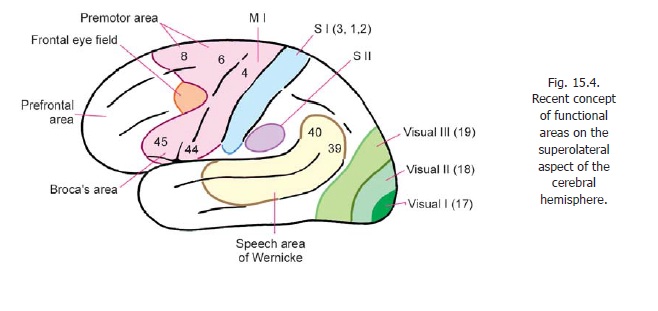
As described the motor area lies in the precentral gyrus (area 4). This is theprimarymotor area (MI). This area has the lowest threshold of stimulation for producing contralateral bodymovements.
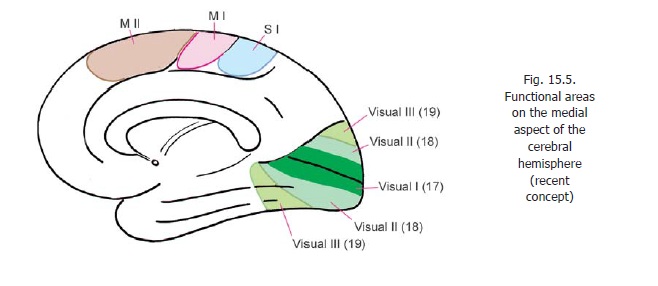
A supplemental motor area (MII) is present on the medial aspect of the cerebral hemisphere, in the medial frontal gyrus, anterior to the paracentral lobule (areas 6, 8). In this area the body is represented with the area for the lower limb posteriorly, that for the face anteriorly, the area for the upper limb lying between these two. Additional motor areas have been described in the adjoining part of the cingulate gyrus.
On the superolateral surface, the premotor area lies anterior to the motor area occupying areas 6 and 8. (We have seen that on the medial aspect of the hemisphere areas 6 and 8 are occupied by the supplemental motor area.)
Advanced:
The distinction between motor and premotor areas is vague, and the entire area is sometimes described as the primary motor area. Studies on areas of the cortex giving origin to corticospinal fibres show that they arise from a wide area covering the main and supplemental motor areas, the premotor area, and many areas in the parietal lobe (SI including areas 3, 2, 1; area 5; and SII in the parietal operculum). It is obvious that this area is not coextensive with the definition of motor areas. Some studies claim that only 20 - 30% of corticospinal fibres arise from area 4; and that as many as 50% may have their origin in the parietal lobe. In relation to motor function the site of termination of corticospinal fibres in spinal grey matter becomes significant. Most fibres that descend from the parietal lobe end in the dorsal grey column; while those ending in the ventral grey column are predominantly from the frontal lobe. The latter appear to be the ones most important for motor control.
The premotor area appears to be responsible for programming intended movements, and control of movements in progress. It is divisible into a dorsal and a ventral area. The dorsal area appears to be concerned with movements initiated by the individual, while the ventral area appears to be concerned with control of movements that take place in response to external stimulation.
The cortex of the motor area is characterised by the presence of large pyramidal cells (giant pyramidal cells of Betz). At one time these cells were considered to be the source of all corticospinal fibres but, as discussed above, this if now known to be incorrect.
It has been estimated that the size of the soma of a neuron is proportional to the total volume of its processes. It follows that neurons having very long axons will have large somata. Purkinje cells that give origin to corticospinal fibres descending to sacral segments of the spinal cord can, therefore, be expected to be among the largest pyramidal cells.
Apart from its corticospinal output, the motor area is connected (in a point to point manner) with the main sensory cortex (SI). This explains why neurons in area 4 are responsive to peripheral stimulation. Area 4 receives afferents from the posterior part of the ventral lateral nucleus of the thalamus. This nucleus receives fibres from cerebellar nuclei. In this way the cerebellum projects to area 4. Area 4 also receives fibres from some other parts of the thalamus, the hypothalamus, and some other parts of the cerebral cortex (including the primary sensory area SI).
The frontal eye field and the motor speech area (of Broca) are parts of the premotor area.
Apart from the motor speech area of Broca there are two other areas concerned with motor control of speech. One of these is located in the temporal and parietal lobes (motor speech area of Wernicke) while the other is located in the supplemental motor area (MII) (Figs. 15.4, 15.5).
Clinical:
In summary note the following:
a) A localised lesion of the primary motor area normally produces contralateral monoplegia. An extensive lesion can cause hemiplegia.
b) Lesions of the premotor area have an adverse effect on skilled movements.
c) Lesions of the frontal eye field result in deviation of both eyes to the side of lesion.
d) Lesions of the motor speech area destroy the ability to speak.
e) Lesions of the prefrontal areas lead to personality changes.
Sensory Areas
The sensory area of classical description is also called the first somatosensory area (SI). It receives projections from the ventral posteromedial and ventral posterolateral nuclei of the thalamus, conveying impulses received through the medial, spinal and trigeminal lemnisci. We have seen that different parts of the body have a definite representation within this area.
Advanced:
It has also been shown that different sensations may be represented in different parts within the area. Unit recordings show that area 2 is concerned mainly with proprioceptive impulses, while area 3 responds only to cutaneous stimuli.
A second area predominantly somatosensory in function (second somatosensory area or SII) has been described in relation to the superior lip of the posterior ramus of the lateral sulcus (Fig. 15.4).
The sensory speech area of Wernicke lies in the posterior part of the superior and middle temporal gyri. It extends into areas 39 and 40 of the parietal lobe. This area is responsible for interpretation of speech.
Parts of the superior parietal lobule (areas 5, 7) help us to recognise shape, size and texture of objects
Like area SI, SII receives fibres from the ventral posterior nucleus of the thalamus. It also receives fibres from SI. Descending fibres from SII reach the spinal cord, the nucleus gracilis and cuneatus and the main trigeminal nucleus. Neurons in SII respond best to intermittent stimulation e.g., vibration. SII may be responsible for perception of pain and temperature.
A small part of the primary sensory area, probably within area 2, is believed to serve as a corticalvestibular area.
Clinical:
The effects of damage to sensory areas are as follows.
i. Damage to the first somatosensory area causes loss of sensation (both exteroceptive and prioprioceptive) from the opposite side of the body.
ii. Damage to the second sensory area may lead to inability to appreciate pain and temperature.
iii. Damage to some areas behind the main sensory area (areas 5, 7) interferes with ability to identify objects by feeling them.
iv. Damage to the area of Wernicke leads to failure to understand speech.
Visual Areas
The classical visual area has been described before. This area receives fibres of the optic radiation. It is also called the striate cortex. In addition to the striate cortex additional areas of cortex responding to visual inputs are described. Area 18 (peristriate area) is the second visual area; and area 19 (parastriate area) is the third visual area.
A modified nomenclature recognising five visual areas has been described as follows.
a) First visual area (V1) in area 17.
b) Second visual area (V2) occupying the greater part of area 18, but not the whole of it.
c) Third visual area (V3) occupying a narrow strip over the anterior part of area 18.
d) Fourth visual area (V4) within area 19.
e) Fifth visual area (V5) at the posterior end of the superior temporal gyrus.
The visual areas give off efferent fibres also. These reach various parts of the cerebral cortex in both hemispheres. In particular, they reach the frontal eye field which is concerned with eye movements. Like other ‘sensory’ areas the visual areas are, therefore, to be regarded as partly motor in function. This view is substantiated by the fact that movements of the eyeballs and head can be produced by stimulation of areas 17 and 18 which constitute an occipital eye field. Efferents from the visual areas also reach the superior colliculus, the pretectal region, and the nuclei of cranial nerves supplying muscles that move the eyeballs. There is physiological evidence of a corticogeniculate projection. Fibres also reach the thalamus (pulvinar).
The total number of neurons to be seen in delimited vertical areas of cortex is remarkably constant in different regions. The cortex of the visual area is an exception and has a much greater density of neurons than other parts of the cortex.
Auditory (acoustic) Areas
The classical area for hearing (primary auditory area, A1) has been described. As in the case of the visual areas it has been shown that fibres of the acoustic radiation end not only in the classical or first auditory area, but extend into some neighbouring areas as well. These include a second acoustic area lying in the superior temporal gyrus anterior to A1. A number of other auditory areas have been described.
Efferent fibres arising in the acoustic areas project to the medial geniculate body, and to the inferior colliculus, and may possibly reach motor nuclei of cranial nerves. Some of these efferents may influence the state of contraction of the stapedius and tensor tympani muscles. The acoustic areas are also connected with other parts of the cerebral cortex.
It may be remembered that the fibres of each lateral lemniscus are bilateral. Hence, the acoustic areas in each cerebral cortex receive fibres from both the right and left cochleae. The close relationship of the acoustic areas to Wernicke’s speech area may be noted. The association is significant in view of the obvious relationship between hearing and speech.
Clinical:
As the auditory areas receive impulses from both sides, a lesion on one side produces only partial loss of hearing. Lesions in the secondary auditory area (area 22) interferes with interpretation of speech (word deafness).
Prefrontal Areas
The part of the frontal lobe excluding the motor and premotor areas is referred to as the prefrontal area. It includes the parts of the frontal gyri anterior to the premotor area, the orbital gyri, most of the medial frontal gyrus, and the anterior part of the gyrus cinguli.
Advanced:
The prefrontal area has numerous connections with the thalamus, the corpus striatum, the limbic system, the hypothalamus, the reticular formation, the cranial nerve nuclei, and the pontine nuclei. These connections suggest that this area is concerned with both somatic and visceral activity. Injury to some parts of this region makes the person more docile, but also negligent and lacking in concentration. The prefrontal area appears to be concerned with normal expression of emotions, and the ability to predict consequences of actions. The medial part of the prefrontal area is associated with auditory and visual functions.
Related Topics