Chapter: Human Neuroanatomy(Fundamental and Clinical): Internal Structure of Cerebellum
Few Details about the Cerebellar Cortex
Further Details about the Cerebellar Cortex
Some details about Purkinje cells
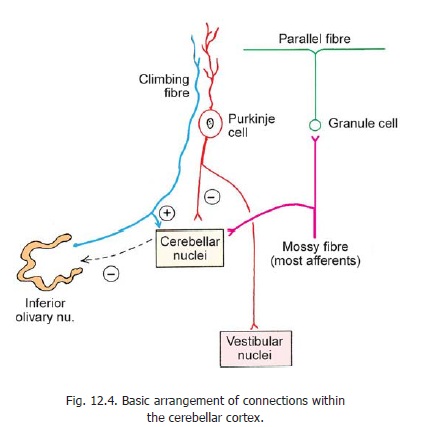
With the EM the dendrites of these cells can be seen to be studded with numerous spines. Some synapses occur on these spines, and others on the ‘smooth’ parts of dendrites between the spines. The dendrites of Purkinje cells synapse with axons of granule cells (parallel fibres), outer stellate cells, and basket cells ; and with climbing fibres (representing olivocerebellar afferents). Climbing fibres synapse with the proximal parts of dendrites while parallel fibres synapse on the distal parts.
The axons give off collaterals that remain within the cortex and synapse with Golgi cells. Some collaterals enter the molecular layer. The part of the Purkinje cell axon next to the cell body resembles the cell body in structure and is called the preaxon.
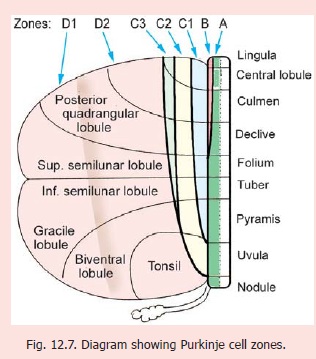
Purkinje cell zones
From the point of view of its connections the cerebellar cortex can be divided into vermal, paravermal and lateral zones. The paravermal zone is also called the pars intermedia. On the basis of recent studies a more detailed map of parasagittal zones (called Purkinje cell zones) has been described (Fig. 12.4). The zones (in each half of the cerebellar cortex) are designated A to D (in medial to lateral sequence). Zones A and B lie in the vermis, zone C in the pars intermedia, and zone D in the lateral area.
Zone A lies immediately lateral to the middleline and includes the whole of the vermis except a strip occupied by zone B .
Zone B occupies the lateral and anterior partof the vermis. It includes the lateral parts of the lingula, central lobule, culmen and declive.
Zone C occupies the pars intermedia. It is divided into three strips C1, C2 and C3 (medial to lateral).Strips C1 and C3 extend caudally up to the level of the pyramis. Strip C2 (lying between C1 and C3) extends further caudally into the medial part of the tonsil.
Zone D is constituted by the part of the cerebellar hemisphere not occupied by zone C. It occupiesthe greater part of the hemisphere. It is sometimes divided into a medial part zone D1, and a lateral part zone D2.
Some details about Golgi neurons
Each Golgi neuron occupies a definite area there being no overlap between the territories of neighbouring Golgi cells. It is interesting to note that the territory of each Golgi cell corresponds to that of about ten Purkinje cells.
The outer stellate cells, the basket cells, and the Golgi cells are inhibitory neurons. Golgi cells are responsible for feedback inhibition of granule cells.
Outer Stellate cells
These cells and their processes are confined to the molecular layer of the cerebellar cortex. Their dendrites (which are few) synapse with parallel fibres (of granule cells) while their axons synapse with dendrites of Purkinje cells (near their origin).
Basket cells
These cell lie in the deeper part of the molecular layer of the cerebellar cortex. Their dendrites that are few ramify in the molecular layer and are intersected by parallel fibres with which they synapse. They also receive recurrent collaterals from Purkinje cells, climbing fibres, and mossy fibres. The axons of these cells branch, and form networks (or baskets) around the cell bodies of Purkinje cells. Their terminations synapse with Purkinje cells at the junction of cell body and axon (preaxon). Each basket cell may synapse with about 70 Purkinje neurons. Basket cells and stellate cells are GABAergic. They are inhibitory to Purkinje cells.
Brush cells (or monodendritic cells)
These are small cells present in the granular layer. Each cell gives off a single dendrite the branches of which give a brush-like appearance. The dendrite synapses with mossy fibres. The axon of the cell is thin and its connections are unknown.
Structure of Glomeruli
The glomeruli are complex synaptic structures. The core of each glomerulus is formed by the expanded termination of a mossy fibre (Fig. 12.8). This termination is called a rosette. Numerous (up to 20) dendrites of granule cells synapse with the rosette. These synapsesare axodendritic and excitatory.
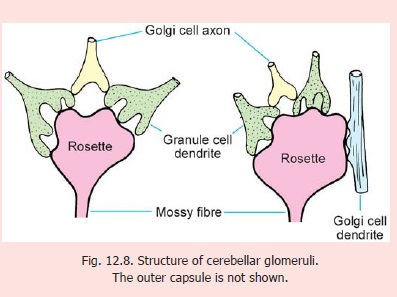
The glomerulus also receives axon terminals of Golgi cells. These also synapse with granule cell dendrites. These synapses are inhibitory. Occasionally, a Golgi cell dendrite may enter a glomerulus and synapse directly with the rosette. The entire glomerulus (which is about 10 μm in diameter) is surrounded by a neuroglial capsule.
Some quantitative data regarding the Cerebellar Cortex
Extensive researches have revealed interesting quantitative data about the cerebellar cortex as follows.
1.The cerebellar cortex has a total surface area of approximately 200,000 square millimeters.
2.The cortex underlying each square millimetre of surface area contains:
(a)about 500 Purkinje neurons;
(b)about 600 basket cells;
(c)about 50 Golgi neurons;
(d)about 3,000,000 granule cells; and
(e)about 600,000 glomeruli.
3.Each axon reaching the cerebellar cortex from the olive divides into about ten climbing fibres. Each olivary neuron, therefore, establishes connections with about ten Purkinje cells.
4.Each mossy fibre synapses with about 400 granule cells. The axons of each granule cell synapse with about 300 to 450 Purkinje neurons.
5.Each Purkinje neuron may bear up to 80,000 synapses with different parallel fibres. The dendritic tree of a single Purkinje cell may be crossed by about 250,000 parallel fibres.
Neurochemistry of the Cerebellum
Recent researches have revealed the presence of several neuroactive substances, and receptors for them, in the cerebellum. Details of these are beyond the scope of this book. Some facts are as follows.
1.Neuroactive substances present in the cerebellum include L-glutamate, aspartate, GABA, serotonin, noradrenalin, acetyl choline, enkephalin and somatostatin.
2.The main neurotransmitter in the cerebellum is L-glutamate. Afferents reaching Purkinje cells through climbing fibres and through the mossy fibre — granule cell — parallel fibre system are glutamatergic. They are excitatory. Some mossy fibres arising in vestibular nuclei are cholinergic. Some reticulocerebellar fibres are serotoninergic, while coeruleo-cerebellar fibres are noradrenergic. Some dopaminergic and cholinergic fibres are also present.
3.Purkinje cells themselves are GABAergic and are inhibitory to cerebellar (and vestibular) nuclei. Some Purkinje cells contain a class of proteins called zebrins. Zebrin containing and non-zebrin containing Purkinje cells may occupy alternating bands of cortex.
Golgi cells, stellate cells and basket cells are GABAergic. Some basket cells, and some granule cells, show the presence of nitrous oxide (NO) which may be a neurotransmitter.
4. Neurons in cerebellar nuclei are glutamatergic (excitatory), or GABAergic (inhibitory). The latter project to olivary nuclei. Some neurons in cerebellar nuclei are glycinergic. These are interneurons and may contain GABA in addition to glycine.
Apart from the neurotransmitters associated with them, cerebellar nerve fibres show the presence of various other neuroactive substances. These include enkephalin, somatostatin, aspartate, and corticotropin releasing factor.
Cerebellar neurons show interesting specialisations of their enzyme systems.
Receptors for neurotransmitters have been demonstrated in cerebellar neurons. Several types of glutamate receptors and GABA receptors can be distinguished. The receptors may be ionotropic (coupled to ion channels) or metabotropic (coupled to other messengers). Some are voltage dependent and cause opening of Ca++ channels in postsynaptic neurons.
Related Topics