Chapter: Introduction to Botany: Growing Diversity of Plants
Bryophyta: the mosses - Growing Diversity of Plants
Bryophyta: the mosses
Bryophyta has
gametophyte predominance while Pteridophyta and Spermato-phyta both have
sporophyte predominance (and the main difference between Pteridophyta and
Spermatophyta is that Spermatophyta has seeds).
Bryophyta
has approximately
20,000 species. They do not have roots, but have long dead cells capable of
water absorbency via apoplastic transport, these cells are called rhizoid cells. Their sporophyte is
reduced to sporogon, which is simply
a spo-rangium with seta (stalk), and
is usually parasitic. Gametophyte of bryophytes starts its development from a protonema, thread of cells. Bryophyta
are poik-ilohydric; they go through dehydration or extremely low water
concentration without any serious physiological damage to the plant.
Bryophyta Life cycle
of mosses is similar to the general life cycle of land plants described above.
They begin with a gametophyte with an archegonia and antheridia. The
antheridium produces biflagellate spermatozoa which fertilizes the egg and
pro-duces diploid zygote; zygote grows into a sporogon and its cells (mother
cells of spores) go through meiosis which produces haploid spores. Spores will
be dis-tributed with the wind, land on the substrate and germinate into
protonema stage which then develops into a green, well-developed gametophyte.
Most of moss gametophytes have a shoot body that consists of a stem and leaves
(but no roots) while others have a thallus body, which is a flat, leaf-like,
and undifferen-tiated structure.
There are three main
groups, also known as subphyla, of Bryophyta: Hepaticae (liverworts),
Bryophytina (true mosses), and Anthocerotophytina (hornworts).
Hepaticae are phylogenetically closest to green algae. Their thallus typicallyhas
dorsal and ventral parts, and the sporogon is bag-like. Inside the sporangium,
there is no central column (columella) but elaters are present, which are cells
that loosen spores. One of the most widespread liverwort is Marchantia, it is commonly found in wet
shady places. It became a frequent weed in greenhouses.
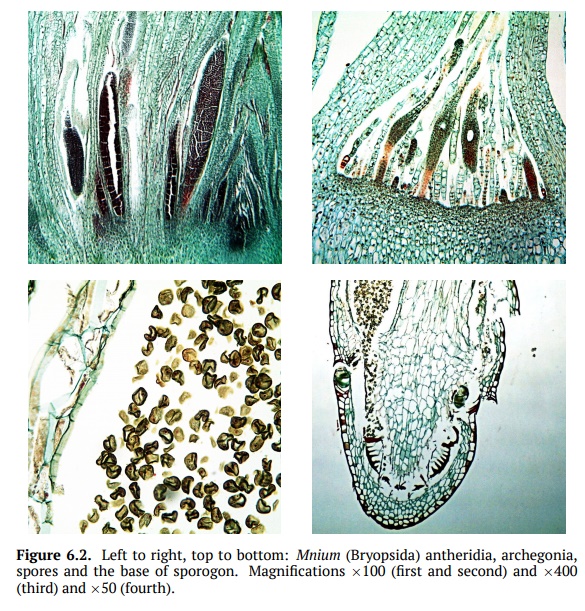
Bryophytina consists of multiple classes, the most important are Sphagnop-sida—peat mosses,
Polytrichopsida—hair cap mosses, and
Bryopsida—greenmosses (Fig. 6.2).
Bryophytina have a radially structured shoot-like body with a stem and thin
leaves. Their sporogon is long and has columella, but does not have elaters.
Sporogons of true mosses are usually supplied with peristome, structure which helps in spore distribution. Some
advanced true mosses (hair cap moss, Polytrichum)
have tall gametophyte with proto-vascular tissues, while others (stinkmoss, Splachnum) employ insects for the
distribution of spores. Peat moss (Sphagnum)
is probably the most economically important genus of Bryophyta.
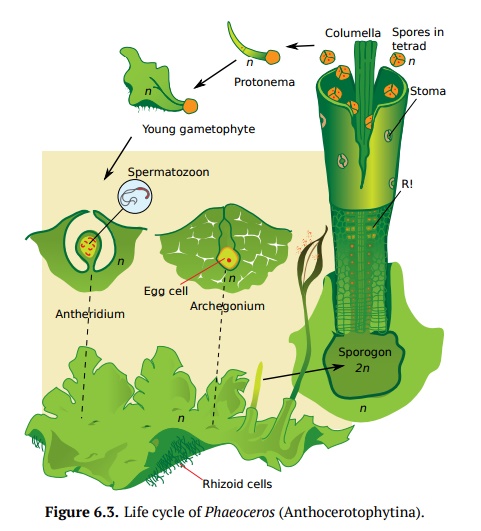
Anthocerotophytina (Fig.6.3)evolutionary are closests to the
next phylum,Pteridophyta (ferns and allies). Hornworts have a flattened thallus
body, their long photosynthetic sporogon has columella and elaters. The
presence of stom-ata on sporogons and the ability of some hornwort sporogons to
branch and sometimes even live independently from the gametophyte provide a
support for the advanced position of this group. Hornworts are rare and quite
small (first millimeters in size), and like liverworts, they prefer shady and
wet places.
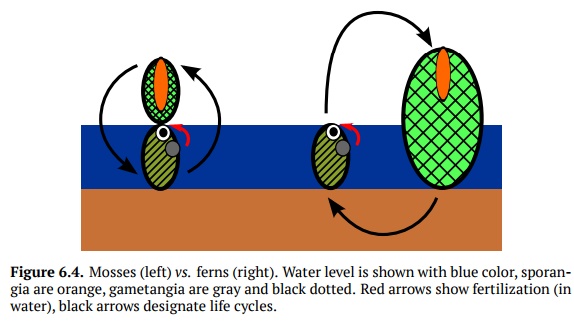
Mosses have become
known as the “evolutionary dead end” because their poik-ilohydric gametophyte
requires water for fertilization and does not have a root system; this
restricts the size and requires dense growing. However, if the sex-ual organs
are near the soil surface, then the parasitic sporogon would not grow tall
enough, and consequently would not be able to effectively distribute spores
with the wind. The only way to fix the situation properly would be to make the
sporophyte taller and reduce dominance of the gametophyte. This is done in
ferns (Fig. 6.4).
Related Topics