Chapter: Plant Anatomy:An Applied Approach: Morphology and tissue systems: the integrated plant body
Adaptation to aerial growth
Adaptation to aerial growth
To understand the structure – morphology and anatomy – of land plants we have to remember that plant life started from single-celled organisms in an aquatic environment. There are still many thousands of different species of unicellular algae both in water and exposed – on tree trunks, leaves, soil and rock faces for example, in suitably moist places. Evolution of algae in the water has produced some very large, multicellular forms, for example Lami-naria species, kelps. These large plants are fine in water, but lack the adapta-tions necessary for terrestrial life. They need to be bathed in water, which is a source of dissolved nutrients. Because they can absorb nutrients over most of their surface area, there is no need for a complex internal plumbing sys-tem, like the xylem (woody tissue) and phloem (cells adapted to conduct synthesized materials in the plant) in vascular bundles of land plants. They lack roots, but have holdfasts, structures adapted to anchor them to a firm substrate, but which are not absorbing organs for minerals and water, such as roots usually are. They lack a waterproof covering, a modified outer layer of epidermal cells of land plants, and rapidly desiccate if exposed to the air. Their mechanical support comes from the surrounding water, so they do not need the woody tissue (xylem) or fibres (elongate, thick-walled cells with tapered ends whose cell walls become strengthened with lignin, a hard material, at maturity; form part of the sclerenchyma) of land plants. True, they are tough and very flexible, and most can survive violent wave action. Even their reproduction depends on the release of male and female gametes into the water around them.
Some types of land plants still rely on a film of water for their male gametes to swim in to reach the female gamete and effect fertilization, for example mosses and ferns, but the higher plants like gymnosperms and angiosperms have their male gametes delivered in a protective package, the pollen grain, to a receptive female part of the cone or flower.
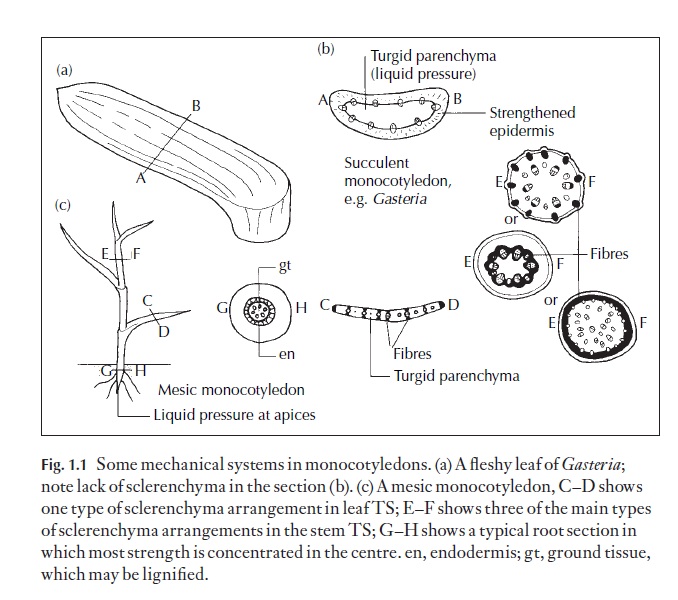
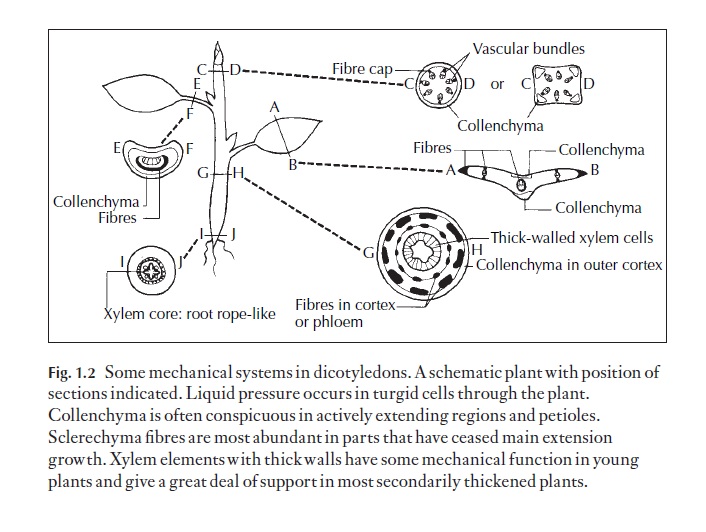
There is a very wide range of land habitats, and land plants show a re-markable range of shapes and sizes. Anatomy of flowering plants (angiosperms), and the vast majority of these share distinct vegetative organs that are readily recognized. They are leaf, root and stem (Figs 1.1, 1.2). These organs cope with the need to obtain, trans-port and retain enough water to help prevent wilting, carry dissolves minerals and keep the plants cool when necessary. Most land plants contain specialized cells and tissues for mechanical support and others for movement within the plant of materials they synthesize. The tough skin (epidermis, together with a cuticle and sometimes waxy materials) pre-vents water loss but permits gas exchange. Small pores in the epidermis of most leaves and young stems can be opened and closed and regulated in size. These are called stomata and they regulate the rate of movement of water and dissolved minerals through and out of the plant. Sometimes the epidermis is the main part of the mechanical system as well, and holds the main leaf or stem material inside under hydraulic pressure.
In many plants, the strength of the ‘skin’ is supplemented by tough me-chanical cells arranged in mechanically appropriate areas. These are forms of sclerenchyma cells with lignified walls: fibres which are elongated cells and sclereids, which are usually relatively short; a range of types exists (see the Glossary). Collenchyma is also a supporting or mechanical tissue which occurs in young organs and in certain leaves; the walls are mainly cellulosic. Here walls are thickest in the angles between the cell walls, or in lamellar collenchyma wall thickening is found mainly on anticlinal cell walls; see below for details.
Plants submerged in water are afforded some protection from damaging ultraviolet (UV) light. Land plants need other mechanisms to prevent UV damage. The green pigment, chlorophyll, is readily damaged by UV. Since this pigment and its cohort of specialized enzymes is responsible for transforming the energy of sunlight through its action on CO2 and H2O into sugars, the starting point for nearly all stored organic energy on earth, it is vitally important that the UV screening methods developed are effective.
All green plants need light for photosynthesis. Plants have evolved differ-ent strategies which bring leaves into a good position for obtaining the sunlight. Some (annuals, ephemerals) put out their leaves before others neighbouring plants, complete their annual or shorter cycle and form seed for the next generation. Others retire to a dormant form (some perennials and biennials) at a time when they may be shaded by taller vegetation. Manyspecies develop long stems or trunks and expose their leaves above the competition (some are annuals or biennials and but most are perennials). Some species do not have mechanically strong stems, but use the support provided by those which do, climbing or scrambling over them (they can be either annuals or perennials). Biennials are plants with a two-year life cycle. They build up a plant body and food reserves in the first year, and then flow-er and fruit in the second.
In summary, the main factors which all terrestrial plants with aerial (above-ground) stems and their associated leaves have to overcome are:
1.Mechanical, i.e. support must be provided in one way or another so that a suitable surface area with cells containing chloroplasts can be exposed to the sunlight to intercept and fix solar energy. These chlorenchyma cells may be on the surface, or just beneath translucent layers of cells. See below for more detail of the cell types that give mechanical strength. Secondary growth in thickness is another strategy that provides mechanical strength to parts both above and below ground. The growth in thickness may be relatively small in annuals, but in perennial plants it may be extensive, and requiring the use of large quantities of energy in its production. When present, the way secondary growth occurs differs between monocots and dicots.
2.Risk of excess water loss, i.e. they must be provided with protection against too much water loss from the exposed surfaces. This is generally done by a combination of a waxy outer layer and a fatty cuticle above an epi-dermis (the outer skin). Because water has to evaporate from some exposed surfaces so that movement of water and dissolved minerals can take place through the plant (transpiration), most leaves, and stems which retain the epidermis, have regulated pores, stomata, which can be opened and closed in response to prevailing conditions.
3.The ability to move water and minerals from the soil (transpiration) through the roots to regions where they can be combined with other mate-rials to build the plant body, and the movement of synthesized food material from the site of synthesis to places of growth or storage and from the stores to growing cells (translocation). Of particular interest is the level of struc-tural and physiological control of the phloem loading process. Epiphytes are attached by their roots to other plants, and obtain their water and minerals in different ways.
4.Reproduction, placement of reproductive organs enabling the pollen or gamete receptor mechanism to operate successfully, and after fertilization and spore/seed production, ensuring dispersal of the propagules.
The first three issues outlined above are dealt with by well-organized (if complex) systems in the higher plants, and will be summarized here.
Related Topics