Chapter: Plant Biology : Plants, water and mineral nutrition
Water retention and stomata
WATER RETENTION AND STOMATA
Key Notes
Plant types
Plants are categorized as mesophytes adapted to an environment in which water is available, xerophytes adapted for areas of low water availability and hydrophytes adapted to the presence of large amounts of water or to growth entirely in water.
Plant structure and surfaces
Leaf structure is adapted to regulate and minimize water loss in unfavorable conditions. The close-packed, flattened epidermal cells of leaf surfaces are covered in hydrophobic cuticles made of cutin and wax, which minimize evaporation. Stomata are most abundant on the lower side of the leaf and may be sunken into the leaf surface.
Stomata: structure
Diffusion of gas is essential for photosynthesis. Stomata (singular, stoma) permit gas diffusion into the leaf while minimizing and regulating water loss. The stomatal pore penetrates the epidermis into the substomatal cavity in the mesophyll. The pore is bounded by two guard cells, which are associated with subsidiary cells in the epidermis. Stomata are open when the guard cells are turgid and closed when they lose turgor.
Stomata: action and regulation
Stomatal opening responds to a number of environmental factors, in addition to drought, including: CO2 concentration, light and temperature. Stomata are held open by turgor from high solute concentrations in guard cells. Abscisic acid causes release of solutes via regulated anion channels in the plasma membrane. This reduces the osmotic potential of the guard cell and water flows from the cells. Opening results when the anion channels close and solutes re-enter the cell.
Stomatal adaptations of xerophytes
Xerophytes are adapted to low water availability. Typical characteristics include: thick fleshy leaves or no leaves (succulence), protected stomata and thick cuticles. Plants showing crassulacean acid metabolism (CAM) open their stomata at night to fix CO2 as malate which is stored in the vacuole, while they remain closed in daytime when evaporative losses are greatest.
Plant types
Mesophytes are adapted to an environment in which water is generally available in the soil and can be extracted by transpiration to supply the needs of the plant. Mesophytes can restrict water loss by the closure of stomata, but desiccation to below 30% total water content normally results in death. Mesophytes may show adaptations to either drought or waterlogging to different degrees depending on species. Some, like corn, form aerenchyma in response to waterlogging, while in others (e.g. rice) it is always present . Presence of thicker cuticles, fleshier leaves, sunken stomata and photosynthetic modification indicate increasing adaptation to drought conditions. Other adaptations to drought include drought avoidance, for instance over-wintering as seeds and loss of leaves or other aerial parts.
Xerophytes are adapted to environments in which water is generally very scarce. They display a range of adaptations, including: sunken stomata(that entrap a layer of unstirred air); stomata open at night; modified photosynthetic mechanisms ; thick cuticles and succulence (thick, fleshy leaves or the absence of leaves and presence of fleshy, modified stems). Cacti and some euphorbias are examples of xerophytes.
Hydrophytes are adapted to live submerged or partially submerged in water. They display modified leaves and stems, and frequently contain air-spaces (aerenchyma) to supply oxygen to underwater organs.
Plant structure and surfaces
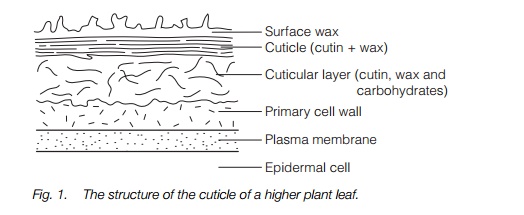
Land plants face a dilemma; how to acquire sufficient carbon dioxide and light for photosynthesis without losing large quantities of water by exposing large evaporative surfaces to the atmosphere. In xerophytes, the area exposed is reduced and lower growth rates occur. Mesophytes with a large leaf area also show adaptations to minimize water loss. Surfaces are coated with a cuticle of lipids, cutin, suberin and waxes which both reduces water loss and the access of pathogens to the cells of the leaf.
Cutin is a polymer of long-chain fatty acids forming a rigid mesh through ester linkages. Suberin contains long-chain fatty acids joined through ester linkages with dicarboxylic acids and phenolics. It is the predominant coating of underground parts of the plant and is also found in the Casparian strip of the endodermis . Associated with cutin and suberin are waxes, longchain acyl lipids which are solid at ambient temperatures. The cuticle is deposited in layers (Fig. 1); first a cuticular layer of cutin, wax and carbohydrates at the epidermal cell wall, then a layer of cutin and wax and finally a waxy surface layer exposed to the atmosphere.
Stomata: structure
Stomata are pores through which gas exchange to the leaf takes place. The pore is formed by two specialized cells, the guard cells, that open and close the pore, frequently associated with subsidiary cells (Fig. 2). The pore structure leads to the sub-stomatal cavity surrounded by cells of thespongy mesophyll. The stomata may themselves be sunken in order to minimize evaporative air movement.
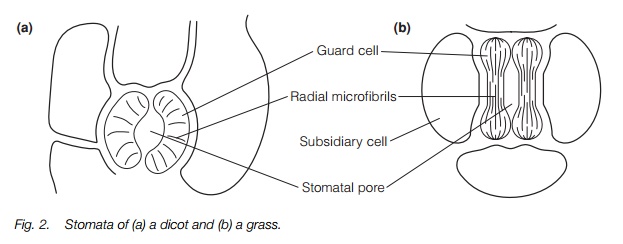
Stomata: action and regulation
Stomatal aperture is tightly regulated by the plant, in a system, which integrates carbon dioxide requirement with light and water stress. Stomatal aperture often varies according to a circadian (day/night) rhythm. Low CO2 concentrations in the guard cells result in opening, while high CO2concentrations result in closure. Stomata in most species open at dawn and remain open in daylight, given adequate water availability; this may in part be due to changing CO2 concentrations as a result of photosynthesis and respiration.
Stomatal guard cells are also very sensitive to water stress. Localized turgor loss results in wilting of the guard cells (hydro-passive closure) and stomatalclosure. Water stress elsewhere in the plant results in the production of abscisic acid (ABA, Topic F2) which results in stomatal closure. The mechanism of ABAsensing probably involves ABA receptor proteins and may involvethe action of calcium as an intracellular messenger that alters membrane ionchannel activity, giving altered turgor and stomatal closure.
Guard cells take up K+ and increase in turgor in favorable conditions, resulting in swelling of the cells and opening of the pore. Potassium is rapidly lost from the guard cells during stomatal closure. The driving force for potassium uptake is provided by the plasma membrane proton pump . This is stimulated during stomatal opening and the resulting membrane hyperpolarization is believed to open K+ channels in the plasma membrane which permit a passive K+ influx. This, together with influx of Cl– and organic anions such as malate provides the increase in turgor. When stomata close, other plasma membrane ion channels open resulting in a rapid efflux of anions and a drop in turgor.
Stomatal adaptations of xerophytes
Xerophytes show a number of adaptations to water stress, including sunken stomata, thickened cuticles and succulence. A key adaptation is the presence of crassulacean acid metabolism (CAM; Topic H3). CAM plants show a specialized rhythm of stomatal action that minimizes water loss. Coupled with their unique metabolism their stomata only open at night, when they fix CO2 as malate in the vacuole. The stomata then remain closed during the day, when evaporative losses would be greatest.
Related Topics