Chapter: Plant Anatomy:An Applied Approach: Meristems and meristematic growth
Practical applications and uses of meristems
Practical applications and uses of meristems
The special properties of the simple, thin-walled meristematic cells, whose anatomy and location in the plant are described above, make possible a number of horticultural techniques. Understanding the position of the meristems in the whole plant, and the delicate nature of meristematic cells, is of importance to those who wish to increase plants by vegetative propaga-tion, increase the number of branches in a plant, make grafts, and improve the chances of good wound healing.
Apical meristems
The main practical uses for apical meristems, particularly shoot meristems, is meristem culture so that new plants can be produced vegetatively. The cells of meristems are undifferentiated parenchyma, and are in an ideal state for cell division to take place. As soon as the meristems are removed from the plant, their formal organization appears to be disrupted, and they are vulnerable to desiccation. They must be cut off carefully and trans-ferred immediately to a nutrient medium. All stages of this technique must be aseptic so that no pathogens are introduced. If the process is successful the apex will first form a mass of callus-like tissue, similar to the orchid protocorm. Small embryonic shoots and roots form subsequently. If the tissue mass is subdivided, a number of small plantlets can be produced. It is important to have a correctly formulated growth medium often particular to the plant under culture or the tissue mass may produce only shoots or roots!
There are several circumstances where it is desirable to reproduce plants by meristem culture. For example, the required plant may be infertile, as inthe case of a triploid, or it could be an F1 hybrid that would not breed true. It is also a useful method for the rapid increase of nursery stock for commer-cial purposes. Other vegetative methods of propagation might take several additional years before a similar number of plants could be produced. Virus diseases rarely infect apices, and meristem culture can be used to produce virus-free stock from otherwise infected plants, for example in the raspber-ry and the potato. Species nearing extinction may be rescued and multi-plied by meristem culture. This may be the only practical approach if the breeding population is very small, or if those plants remaining are self-incompatible. Unfortunately, all of the products of an individual meristem culture will have the same genotype, so genetic diversity cannot be increased by this method of multiplication.
![]()
![]() As a method of propagation, meristem culture would appear to have a bright future. It probably has more potential than the longer established callus culture method, whereby small portions of excised tissue (usually parenchyma) from various parts of a plant are cultured in, or on, a nutritive medium. It may take a long time to induce embryonic plants to differentiate from such a callus.
As a method of propagation, meristem culture would appear to have a bright future. It probably has more potential than the longer established callus culture method, whereby small portions of excised tissue (usually parenchyma) from various parts of a plant are cultured in, or on, a nutritive medium. It may take a long time to induce embryonic plants to differentiate from such a callus.
When large enough to handle, the embryonic plants are detached and grown on a sterile medium to a size at which they can be potted on, in nor-mal potting compost.
Intercalary meristems
Intercalary meristems are also used in horticulture for propagation. In the plant one of their functions is to cause a stem that has fallen over to grow back in an upright position, for example in Triticum, carnation. Carnations will serve as a practical example of where an intercalary meristem is capable of producing adventitious roots. In Fig. 2.7 a carnation stem is shown cut off the plant just below a node. It is split longitudinally through the node, into the intercalary meristem zone. In horticultural practice, the split is often held open with a small piece of stick, until adventitious roots develop from the split sides.
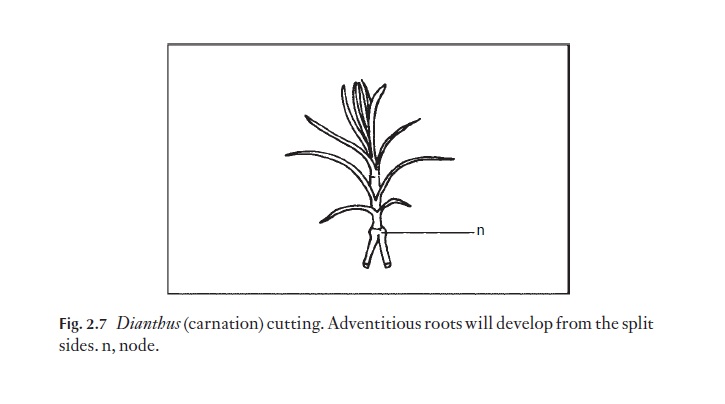
A large number of plants quite readily form adventitious roots from the nodes, whether split off the plant or not. Considerable use is made of this property in horticulture for propagation.
Lateral meristems
Lateral meristems are also used in techniques designed to propagate plants by cuttings and in grafting, or in promoting wound healing.
The cork cambium is so specialized as to be of little value in plant propa-gation. It frequently plays a part in wound healing, and is, of course, em-ployed commercially in the production of ‘cork’ fromQuercus suber, the
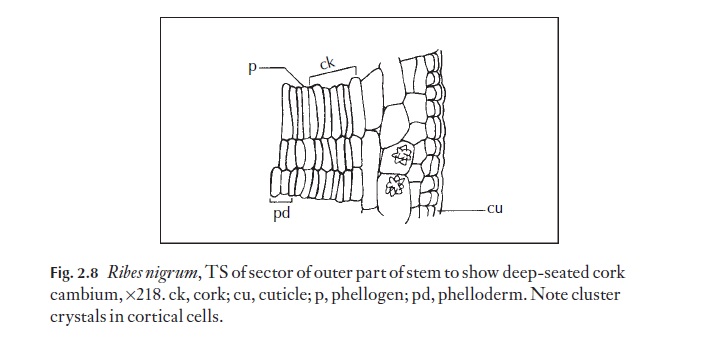
cork oak, in which the cork layers are harvested at approximately ten-year intervals. Cork cambium or phellogen re-forms after the cork is carefully removed. Figure 2.8 shows a cork cambium in Ribes nigrum.
Of the lateral meristems, it is the vascular cambium between phloem and xylem that is most often employed by horticulturists. Its normal function in the healthy woody or herbaceous dicotyledon is to produce new cells of phloem and xylem.
If a cambium is wounded, it will normally regenerate and by influencing the developmental pathways of callus cells next to it, will assist in the heal-ing process, so that cambial continuity is regained, and new cylinders of phloem and xylem established.
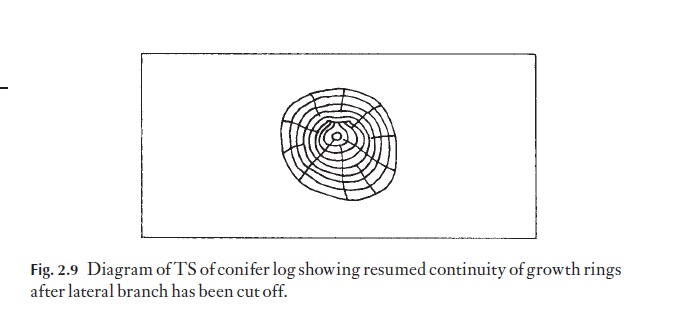
The forestry practice of removing lower branches on conifers at an early stage enables the wound to heal over (Fig. 2.9) and entire rings of sound new wood may become established. If ‘snags’ or broken ends of branches are left, it is some time before they are grown over, and bad knots of dead tissue and hence weak places in the harvested timber will inevitably result.
The inherent ability of wounds to heal is widely used in grafting tech-niques. In order to establish a graft, the parts of the two plants to be united are ‘wounded’. This is done by cutting the root stock and scion (short sec-tion of stem, with buds, or bud alone). The two are brought together in such a way that the cambia of stock and scion are aligned as closely as possible. When new growth is formed by callus cells, the two cambia can then quick-ly establish continuity by specialized differentiation of some of the callus cells and a firm, even bond is produced. No cell fusion takes place, but even-tually the xylem products of the two (joined) cambia firmly bond stock and scion together. It is essential that the stock and scion should not be able to move relative to one another during the early stages, and in these stages grafting tapes are used which both give a secure bond and permit diffusion of oxygen essential to cell growth. The tape must either perish by itself in due course, or be easily removed when the graft is secure. Aftercare is very expensive for the horticulturist, and the simpler the method and the less handling involved, the better. Air gaps between stock and scion must be avoided; they can harbour pathogens, or permit the entry of water and pathogens.
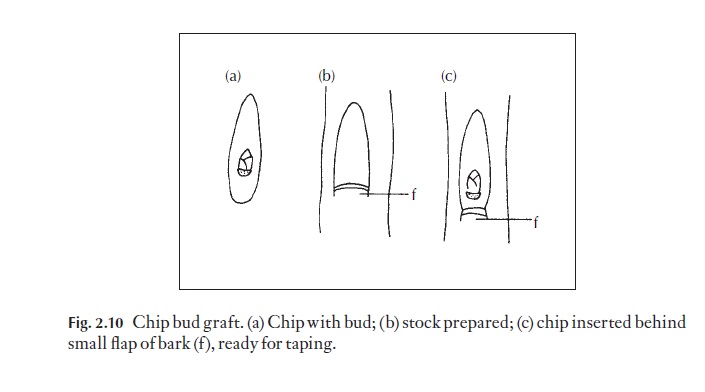
Bud grafting works in much the same way, and the chip bud graft (Fig. 2.10) is becoming increasingly popular and replacing the older T-cut method. The bud cambium can be aligned more accurately by this newer method. A chip bud is removed and inserted behind the small lower lip of bark of the stock at the depth of the cambium and the bud secured by grafting tape.
The advantages of grafting are manifold. For instance the roots of some desirable species may be very weak, and vigorous roots can be grafted in their place, as in Juniperus virginiana where J. glauca stock is employed. Water melon with wilt-prone roots can be grafted onto a gourd root stock that is Verticillicum wilt resistant. The sizes of fruit trees, particularly apples and pears, may be regulated by careful selection of root stock vigour. Trees of relatively fixed mature size can be produced, and earlier fruiting induced
In the UK, the Malling Merton system provides trees with numbered root-stocks guaranteeing a mature tree with specific characteristics. Uniformity of size is essential for good husbandry. The dwarfing stocks have xylem ves-sel elements that are much narrower in diameter than those of the stocks producing large trees.
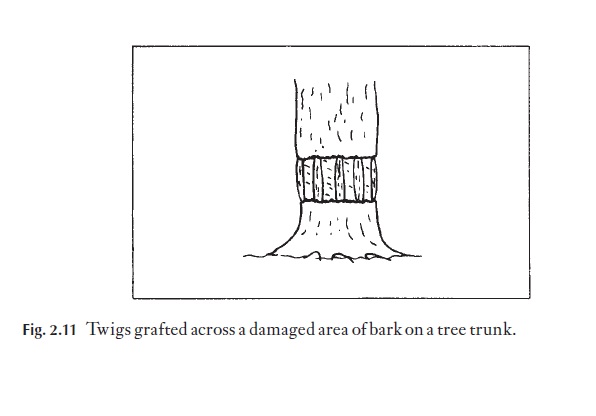
Bridge grafts can be used to repair trees that have been ring barked (Fig. 2.11). It is important to use twigs from the same species, because compati-bility between stock and scion is essential. The twigs must be inserted so that their distal ends point away from the roots, to retain the correct polari-ty. In fact, the interrelationship between plants can be tested to a limited extent by their intergrafting ability. Species from the same genus will fre-quently unite, for example, Prunus species. Solanum species can also be grafted together. Graft hybrids between genera are much less common, for example Laburnum/Cytisus. Grafts between plants from different families occur rarely. However, it has been possible to demonstrate the relationship between the Cactaceae and the Madagascan endemic family Didieraceae from the production of successful interfamily grafts. In the wild, it is quite common for roots of individual trees of the same species growing closely together to become grafted together. Roots abraded by the soil, or damaged by other organisms, form callus, and are thus ‘prepared’ for grafting. The rapid spread of Dutch elm disease between trees in hedgerows is thought to have been due in part to root grafting between individuals.
Bud grafts are used to propagate material rapidly, for example to bring a new rose onto the market quickly. Roses, particularly hybrid teas and flori-bundas, are often poor performers on their own roots, and of course will not come true to type from seed. In such instances, grafting onto a healthy vig-orous rootstock performs the dual function of providing a vigorous root and helps in rapid propagation.
If the vigour of the scion greatly exceeds that of the stock, ugly over-growth can occur, and where there is no desire to regulate scion vigour, a stock of suitable vigour should be selected.
The callus cells produced by wounding two (or more) plants can some-times be grown in culture, and groups of cells centrifuged together. The re-sulting complex of cells can be grown on, producing cytohybrid plants of the most complex type of graft imaginable, that is, except for the fusion of protoplasts of two different organisms that represents the extreme form of grafting! This latter process involves the enzymatic removal of the cell walls, leaving naked protoplasts that fuse more readily.
Monocotyledons are virtually impossible to graft, although there are a few questionable reports of such grafts in the literature. Most monocots have no secondary growth in thickness. The vascular bundles are ‘closed’ and produce no cambium. Some appear to have cambial division, but this may merely be the late, rather regular divisions of cell layers that take place in the central region of the bundle when it is approaching maturity. How-ever, not enough is known about this phenomenon, and it remains ques-tionable whether grafts can become established in monocotyledons.
The monocotyledonous bundle is often firmly enclosed by a scleren-chyma or parenchyma sheath, or both; it is termed a ‘closed’ bundle for this reason. So the monocotyledonous bundle lacks the meristematic cells needed to effect fusion, and the accurate positioning of bundles in a graft would, furthermore, be virtually impossible.
As mentioned earlier, secondary growth in thickness does occur in some monocotyledons, but special tissues at the periphery of the stem bring it about. These are in effect a lateral meristem and produce both new, entire vascular bundles by cell division and the new ground tissue between the bundles. Cordyline has the type of secondary thickening common to a number of monocotyledons. Again it is easy to see that grafting would fail![]()
![]() because it is not possible to align enough bundles and little or no vascular continuity can be achieved.
because it is not possible to align enough bundles and little or no vascular continuity can be achieved.
Adventitious buds
Some plants have the ability to produce new, adventitious buds from various organs when the plant or part of the plant is placed under some unusual physiological stress. The stress may be caused by an injury or even by the separation of one organ from the rest of the plant. The development of buds in this way is usually thought to be related to the loss of a constraint, for ex-ample, the loss of some inhibitory hormone or similar chemical substance. When the apical dominance of a shoot system is removed, new adventitious buds (not related to leaf axis) may develop. This enables us to lop certain species of mature tree and get new growth. For example, Salix and Platanus will grow new branches from adventitious buds. The practice of pollarding and harvesting the pole-like young branches would not be possible if this type of recovery did not take place. Some crops such as Quillaja and Cincho-na bark are obtained from coppiced trees; extracts from these are used inthe preparation of medicines. Much fuel wood production results from pol-larding or coppicing trees. For example, hazel (Corylus) coppice has been a traditional form of husbandry for centuries for forming new stems for char-coal production.
Related Topics