Chapter: Plant Anatomy:An Applied Approach: Adaptive features
Xerophytes
Xerophytes
Plants growing under very dry conditions normally show a reduction in evaporating surface area. When leaves are developed they may be small, or have various features which would appear to assist them in regulating or re-ducing potential water loss. Leafless plants, for example many Cactaceae and Euphorbiaceae, and others with non-functional leaves, for example many Juncus species and most Restionaceae (a family present mainly in the southern hemisphere, in low rainfall areas with mineral-poor soils in Africa and Australasia, and one species in South America), often have subspherical or more or less cylindrical stems modified to perform the photosynthetic and transpirational functions normally ascribed to leaves. A sphere has the smallest surface area possible for a given volume and cylinders also have a low ratio of surface area to volume.
Xerophytic plants may be divided into two categories, drought escapers and drought resisters. Only those that resist drought might be expected to have strongly modified anatomy, but even the escapers that survive as seed or bulbs or in reduced, leafless forms may have some adaptations for the semi-arid periods in which they are normally leafy.
The bulbous habit is often related to dry situations; flowers and leaves are present as aerial organs for a limited period each year, for example Narcissus, Tulipa, Haemanthus,Scilla. Swollen underground stems also occur, for ex-ample many Asclepiadaceae; rhizomes, for example Iris species; or corms, for example Crocus, Watsonia; woody xylopodia, in for example many Australian Anacardiaceae and Myrtaceae. As in ephemerals, these plants normally grow actively when water is available, and their leaves in conse-quence may show little adaptation to xeric conditions. If they are ever-greens, like eucalyptus species, their leaves become tough and show xeric modifications.
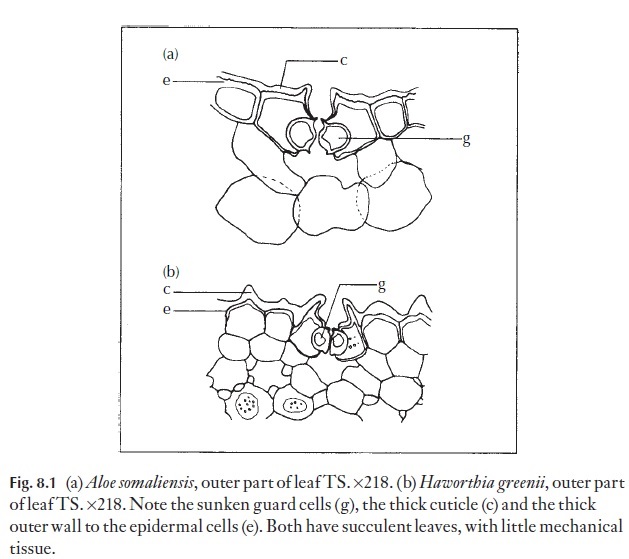
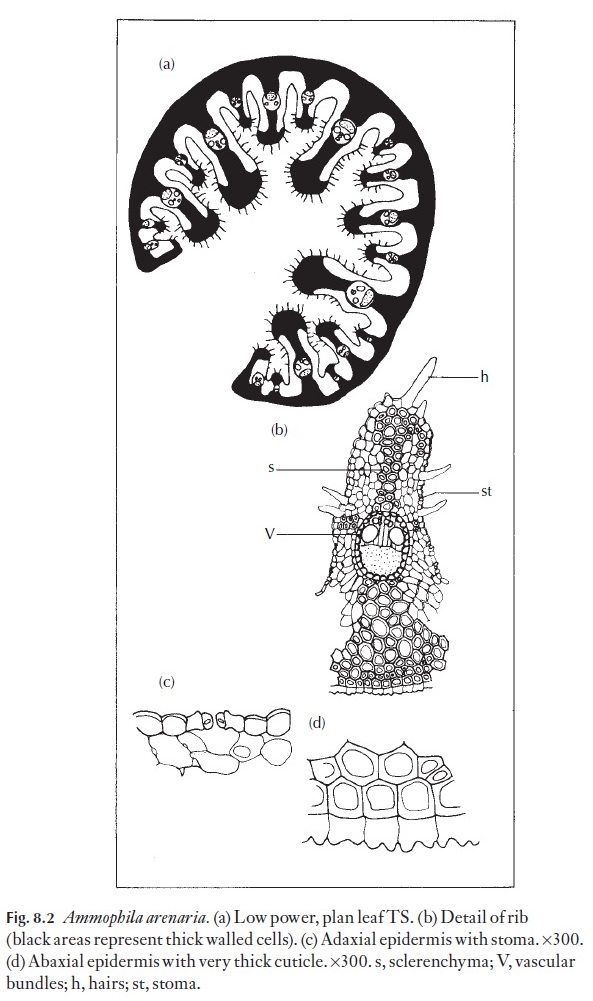
In those plants which have persistent (perennial) leaves or stems, mor-phological and anatomical modifications are quite common. Stomata are often (but not always) sunken; they can have various antechambers and cutin-lined substomatal cavities which could play a part in water regula-tion. Certain Aloe and Haworthia species (Fig. 8.1) show some such modifi-cations. The cuticle itself is often thicker in xerophytes than in mesophytes, but cuticle and epidermal cell wall thickness are not reliable guides to xero-morphy. Stomata may be very numerous and widely distributed, or they may be confined to grooves or channels in the leaf or stem. Some xeromor-phic leaves are capable of inrolling (e.g. Ammophila, Fig. 8.2) and thus en-closing the stomata when dry conditions prevail. On the other hand, when adequate water is available, it has been shown that conifers with needle-like leaves can transpire as rapidly as mesophytes. Plants like many aloes, with thick cuticle, epicuticular waxes, and thick outer walls to the epidermal
cells, sunken and variously protected stomata seem to be quite able to regu-late and minimize water losses during dry periods. The raised rim forming a suprastomatal cavity above each stoma may have a function enhancing evaporation when growth conditions are good. The structure could have a venturi effect, lowering pressure above the stoma and assisting transpira-tion under suitable conditions. In plants like these that have extensive pro-tection from water loss, it may be that even when water is in adequate supply it is difficult to maintain enough evaporation from the leaves to drive the transpiration stream. In these circumstances, if the stomata at the bottom of the chimneys are open, external air flow above the chimneys could cause reduced pressure in them, enhancing the flow of water vapour from the plant.
Some plants, for example Elegia (Restionaceae), can grow in areas of ade-quate groundwater supply, but where strong drying winds could cause ex-cessive water loss. These rush-like plants are not damaged physically by the strong winds since they are leafless and flexible. Many of the Restionaceae and some of the Juncaceae are remarkable in showing xeromorphic features in the stems but hydromorphic features in the roots. There is abundant mechanical tissue, usually sclerenchyma or other lignified cells, in the stems, but large air cavities in the cortex of the roots. It seems that the stems can be exposed to strong drying winds when it is probably too cold for the roots to deliver enough water to meet evaporation losses. The roots themselves often grow in waterlogged soils or standing water. Con-sequently, when conditions for transpiration and root action are satis-factory, the root adaptation is probably beneficial.
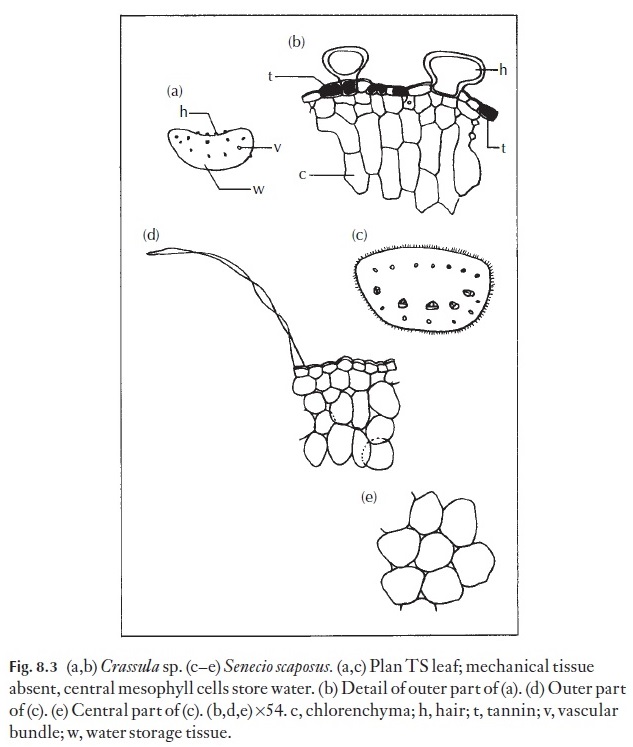
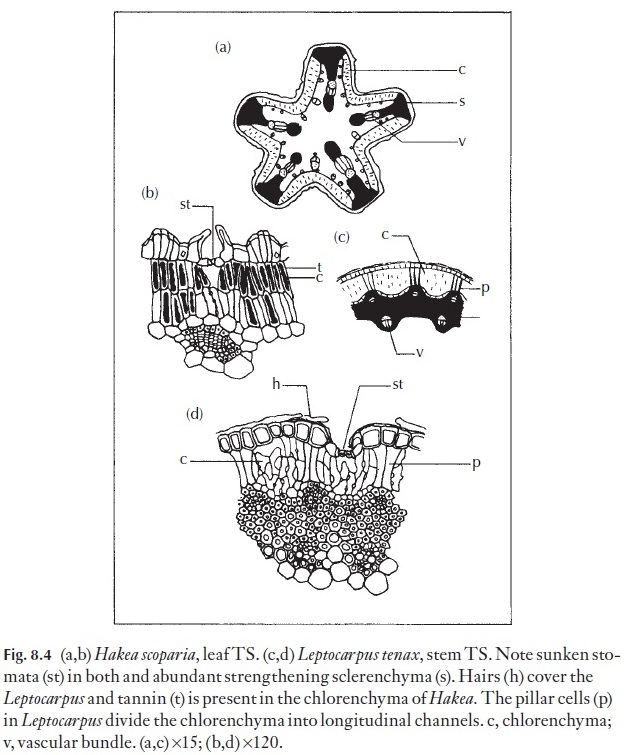
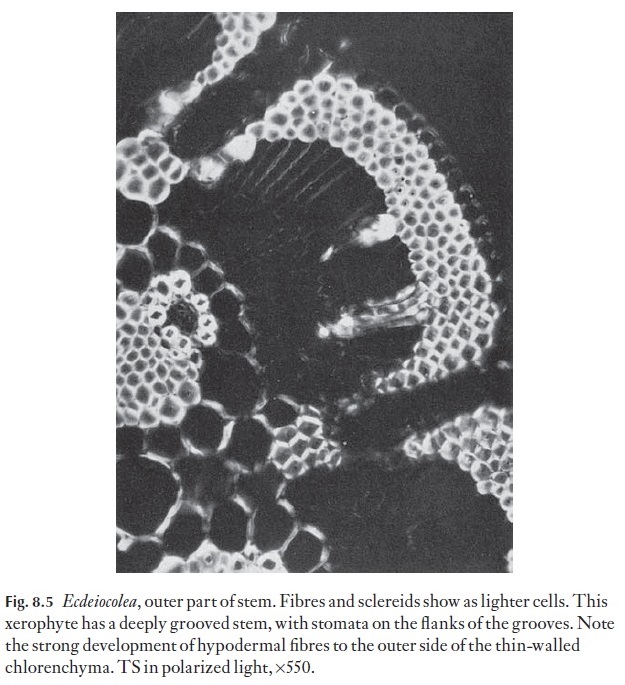
Internal adaptations in xerophytes may take one of two main forms: they can store water, and the plants are then described as being succulent, or they can provide structural rigidity, with the ability to resist collapse and tearing on drying, and then the plants might be described as sclerotic. Tearing and disruption of tissues is one of the main causes of permanent injury resulting from excessive desiccation. Chlorenchyma enclosed in rigid, lined chan-nels is less likely to be torn than that which is in an unprotected, relatively unstiffened mesophyte leaf. In succulent plants there is very little, if any, mechanical tissue, and the xylem of the vascular system is usually not strongly thickened. Many crassulas (Fig. 8.3), aloes, etc. are of the succulent type, and Hakea, Leptocarpus (Fig. 8.4) and Ulex, gorse, are of the sclerotic type, as is Ecdeiocolea (Fig. 8.5).
Some Haworthia and Lithops species show only translucent leaf tips above the ground level. The rest of the leaf is buried, but contains chlorenchyma and water-storing mesophyll. These are often called ‘window’ plants. Light is able to penetrate to the photosynthetic tissue through the translucent cells which function like optical fibres.
The association of some other characters with xerophytes is much more dubious. Hairs have been supposed to help reduce surface wind speed and hence evaporation rates, but hairiness is often much more of a family char-acter. There are many xerophytes belonging to families in which hairs are rare that manage quite adequately without them. Thin-walled hairs could easily increase water loss under some conditions, but most hairy xerophytes, for example Gahnia species, Ammophila and Erica species, have hairs with thickened walls and some also have thick cuticle. Extreme hairiness is found in the plants of high tropical mountains, subjected to extreme diurnal tem-perature fluctuation. Some plants, such as mesembryanthemums, have balloon-like hairs, which when fully turgid allow gas exchange to occur be-tween the air and the stomata beneath them. If water is short, the hairs part-ly collapse, press against one another, and effectively block gas exchange and water vapour loss. Some hairs that do function as an insulating layer, reducing water loss in dry, windy conditions have a waterproof layer of suberin-like material built into their cell walls near the hair base (like a Casparian thickening). This effectively prevents water loss from the body of the leaf through the hairs which might otherwise leak out by a passive wicking effect. In the other extreme, hair cells might have thin walls, and thus be vulnerable to water loss – but equally capable of taking up water from mist or moist air. Hairs like these are common on epiphytes growing high in the canopy of trees in cool mist forests. The roots of epiphytes serve principally for anchorage; water is taken in by the leaves, aided by hairs like these when they are present.
There are numerous mesophytes with hairs.
The fine texture of the leaf or stem surface can have marked effects on the flow pattern of air above it. The boundary layer is the layer of air immedi-ately next to the surface. It is thinner over a smooth than a rough surface. The boundary layer consists of relatively still air, but surface roughness can lead to turbulence in air flowing over it. Water vapour loss through stomata occurs more rapidly when the boundary layer is thin. Species habitually growing in humid environments, sheltered from the wind, tend to be smooth surfaced. In the more exposed environments, for example, hot, rocky hillsides, it is common to find leaves with quite rough surfaces, the roughness increased by prominent flakes of surface wax.
Increased surface roughness can be brought about by leaf form, prominent veins and corrugated laminas. Often it is contributed to by micro-characters as well. In smooth-leaved plants epidermal cells have flat outer periclinal walls. Even a slight doming of these walls can increase sur-face roughness, and if the walls are developed into papillae, they have a marked effect on the boundary layer. Of course hairs, discussed above, in-troduce a new dimension to surface roughness. The outer walls of epider-mal cells may exhibit fine striations and micropapillae. The size of these features, and their occurrence, which is apparently correlated with plants exposed to wind and heat stress, suggests that they play a part in modifying the boundary layer. They may also increase light and heat scatter.
The edges of leaves, their margins, can be entire, or variously toothed (dentate). Dentations have marked effects on reducing turbulent air flow, and are thought to help maintain the integrity of the leaf margins in strong winds.
On the mountain tops, species in less dense air are subject to high insola-tion and have to contend with very high levels of damaging UV light. Chlo-roplasts can bleach and be rendered ineffective if they intercept too much UV light. It has been found that some species have evolved natural sun screens, often involving polyphenols, filtering the UV. These substances may be in the epidermal cells, or in a layer or two of otherwise colourless cells beneath the epidermis but above the chlorenchyma. While in the cells the substances may be translucent and colourless, but sometimes they are yellow to brown, and could have a direct blocking effect on light, reducing its intensity. There is considerable commercial interest in finding out how these chemicals work, and their structure.
Many xerophytes have a hypodermis, the cells of which are thick-walled (Fig. 8.5).
Compact mesophyll, with few air spaces, has also been thought of as a xeromorphic character (e.g. Pinus species, with plicate mesophyll cells), however, many succulent and sclerotic xerophytes have mesophyll or stem chlorenchyma that has abundant air spaces (e.g. Laxmannia, Hypolaena). Experiments are needed to determine the significance of high volumes of internal atmosphere which can be a feature of both xeromorphic and hy-dromorphic plants. Could the function be the same in both types of plant, because they could each be growing under conditions which permit only a low rate of flow in the transpiration stream?
![]()
![]()
There are certain montane parts of the world where little, if any, surface water can be seen for many months of the year, and where soil water is often frozen. The cushion plant is the characteristic life form in such places. From observation of the compact habit, reduced leaf surface area, short inter-nodes, widely penetrating roots and slow growth rate, it might be thought that the anatomy of all such plants would conform to a xeromorphic type. However, this has not proved to be entirely true. Some species, such as Pyc-nophyllum molle and P. micronatum of the Caryophyllaceae, do show the ex-pected adaptation. They have extreme reduction of the leaf surface area, leaves are closely appressed to the cylindrical stems, and sunken stomata are present only on the adaxial protected surface, amongst papillae. But other species, like Oxalis exigua (Oxalidaceae), have very little apparent xeromorphic modification. Oxalis exigua has hairs and papillae on the leaves, but the stomata are superficial. The chlorenchyma of the leaves is not com-pact. Its leaves are similar to those of the mesic members of the genus. The stem and roots exhibit an interesting modification, probably useful to a plant which has to penetrate the cracks between rocks. There is no interfas-cicular xylem or phloem produced during secondary growth and the vas-cular bundles remain separate. Cambium in the interfascicular regions produces parenchyma. Apparently, the roots and stems can twist and de-form without undue compression of the vascular supply, as seen in many lianas.
Azorella compacta (Umbelliferae) shows anatomical features which ap-pear to be related to the harsh high montane environment. The leaves are small and very shiny (thus reflecting ultraviolet light). Contractile roots are present which help to keep the plant firmly anchored, despite frost heave. The vascular bundles are separate, as in Oxalis exigua. Resin ducts, a family characteristic, are frequent.Anthobryum triandrum (Frankeniaceae) is also well adapted to cold, drying conditions. The leaves are furrowed, with sto-mata confined to the furrows. However, the vascular cylinder is compact, not composed of separate bundles. So, once more we have evidence that some ‘family’ characters can be conserved in much modified and reduced plants, and that various species with diverse anatomy can cope with a par-ticular set of environmental conditions.
Halophytes often show succulence which is normally associated with dry conditions; they grow in saline areas where there is, in effect, a physiologi![]()
![]() cal drought. Although surrounded by water, the roots have to extract it from the soil against a considerable suction force. (This point has application in liquid feeding of plants through a gravel bed in glasshouses. The gravel has to be periodically flushed with salt-free water or the salt concentration be-comes high enough to dehydrate the plants.)
cal drought. Although surrounded by water, the roots have to extract it from the soil against a considerable suction force. (This point has application in liquid feeding of plants through a gravel bed in glasshouses. The gravel has to be periodically flushed with salt-free water or the salt concentration be-comes high enough to dehydrate the plants.)
Plants growing in soils which freeze for part of the year are also subjected to ‘drought’ conditions. Many conifers in such habitats have needle-like leaves and the ability to regulate water flow adequately, both in conditions of adequate and inadequate water supply.
The sap of plants which commonly flower and produce leaves before the snow and frost have gone is often of a mucilaginous nature, and acts as a kind of antifreeze.
Ribbed columnar cacti are perhaps the best example of adaptive changes in morphology and anatomy to meet functional needs in a xeric environ-ment. Their shape as a fluted column allows the stem to contract and ex-pand in response to water loss and uptake without damaging any cells. That is the distance between ridges changes with water content, the ribs are clos-er when water stress is happening and farther apart when water has been ac-cumulated. This flexibility is facilitated by the presence of hypodermal collenchyma. Also the shape allows for direct sunlight for only a brief time each day, thereby avoiding overheating of the tissues while the ribs them-selves maximize reflected light. The spines act as shade along the ribs. The thick layer of epicuticular waxes maximizes reflection of damaging ultravi-olet light. The ribs act like radiators in dissipating heat, as do the spines which are modified leaves. During cool nights the ribs and spines are areas where moisture condenses and drips and/or runs to the roots at the base of the plants. So spines not only can prevent herbivory, but they can also pro-vide shade, act as radiators and help collect moisture.
Related Topics