Chapter: Medical Physiology: Sensory Receptors, Neuronal Circuits for Processing Information
Transmission and Processing of Signals in Neuronal Pools
Transmission and Processing of Signals in Neuronal Pools
The central nervous system is composed of thousands to millions of neuronal pools; some of these contain few neurons, while others have vast numbers. For instance, the entire cerebral cortex could be consid-ered to be a single large neuronal pool. Other neuronal pools include the different basal ganglia and the specific nuclei in the thalamus, cerebellum, mesen-cephalon, pons, and medulla. Also, the entire dorsal gray matter of the spinal cord could be considered one long pool of neurons.
Each neuronal pool has its own special organization that causes it to process signals in its own unique way, thus allowing the total consortium of pools to achieve the multitude of functions of the nervous system. Yet despite their differences in function, the pools also have many similar principles of function, described in the following pages.
Relaying of Signals Through Neuronal Pools
Organization of Neurons for Relaying Signals. Figure 46–9 isa schematic diagram of several neurons in a neuronal pool, showing “input” fibers to the left and “output” fibers to the right. Each input fiber divides hundreds to thousands of times, providing a thousand or more terminal fibrils that spread into a large area in the pool to synapse with dendrites or cell bodies of the neurons in the pool. The dendrites usually also arborize and spread hundreds to thousands of micrometers in the pool.
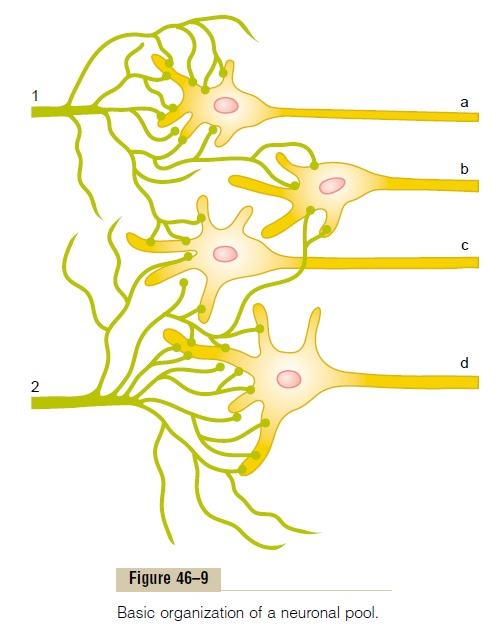
The neuronal area stimulated by each incoming nerve fiber is called its stimulatory field. Note in Figure 46–9 that large numbers of the terminals from each input fiber lie on the nearest neuron in its “field,” but progressively fewer terminals lie on the neurons farther away.
Threshold and Subthreshold Stimuli—Excitation or Facilitation. From the discussion of synaptic function, it will be recalled that discharge of a single excitatory presynaptic terminal almost never causes an action potential in a postsynaptic neuron. Instead, large numbers of input terminals must discharge on the same neuron either simultaneously or in rapid suc-cession to cause excitation. For instance, in Figure 46–9, let us assume that six terminals must discharge almost simultaneously to excite any one of the neurons. If the student counts the number of terminals on each one of the neurons from each input fiber, he or she will see that input fiber 1 has more than enough terminals to cause neuron a to discharge. The stimulus from input fiber 1 to this neuron is said to be an excitatory stimulus; it is also called asuprathreshold stimulus because it is above the threshold required forexcitation.
Input fiber 1 also contributes terminals to neurons b and c, but not enough to cause excitation. Never-theless, discharge of these terminals makes both these neurons more likely to be excited by signals arriving through other incoming nerve fibers. Therefore, the stimuli to these neurons are said to be subthreshold, and the neurons are said to be facilitated.
Similarly, for input fiber 2, the stimulus to neuron d is a suprathreshold stimulus, and the stimuli to neuronsb and c are subthreshold, but facilitating, stimuli.
Figure 46–9 represents a highly condensed version of a neuronal pool because each input nerve fiber usually provides massive numbers of branching termi-nals to hundreds or thousands of neurons in its distri-bution “field,” as shown in Figure 46–10. In the central portion of the field in this figure, designated by the circled area, all the neurons are stimulated by the incoming fiber. Therefore, this is said to be the dis-charge zone of the incoming fiber, also called the excited zone or liminal zone. To each side, the neuronsare facilitated but not excited, and these areas are called the facilitated zone, also called the subthresholdzone or subliminal zone.
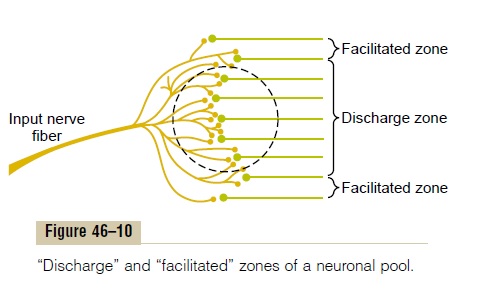
Inhibition of a Neuronal Pool. We must also rememberthat some incoming fibers inhibit neurons, rather than exciting them. This is the opposite of facilitation, and the entire field of the inhibitory branches is called the inhibitory zone. The degree of inhibition in the centerof this zone is great because of large numbers of endings in the center; it becomes progressively less toward its edges.
Divergence of Signals Passing Through Neuronal Pools
Often it is important for weak signals entering a neu-ronal pool to excite far greater numbers of nerve fibers leaving the pool. This phenomenon is called diver-gence. Two major types of divergence occur and haveentirely different purposes.
An amplifying type of divergence is shown in Figure 46–11A. This means simply that an input signal spreads to an increasing number of neurons as it passes through successive orders of neurons in its path. This type of divergence is characteristic of the corticospinal pathway in its control of skeletal muscles, with a single large pyramidal cell in the motor cortex capable, under highly facilitated conditions, of exciting as many as 10,000 muscle fibers.
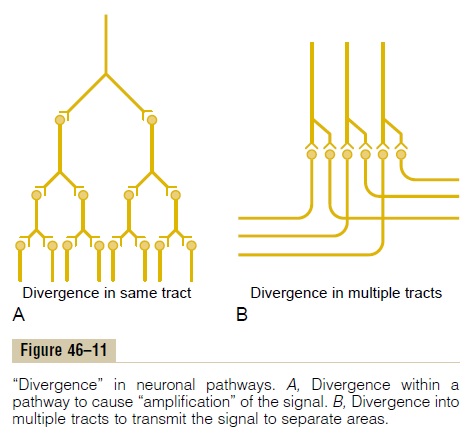
The second type of divergence, shown in Figure 46–11B, is divergence into multiple tracts. In this case, the signal is transmitted in two directions from the pool. For instance, information transmitted up the dorsal columns of the spinal cord takes two courses in the lower part of the brain: (1) into the cerebellum and (2) on through the lower regions of the brain to the thalamus and cerebral cortex. Likewise, in the thala-mus, almost all sensory information is relayed both into still deeper structures of the thalamus and at the same time to discrete regions of the cerebral cortex.
Convergence of Signals
Convergence means signals from multiple inputsuniting to excite a single neuron. Figure 46–12A shows convergence from a single source. That is, multiple ter-minals from a single incoming fiber tract terminate on the same neuron. The importance of this is that neurons are almost never excited by an action poten-tial from a single input terminal. But action potentials converging on the neuron from multiple terminals provide enough spatial summation to bring the neuron to the threshold required for discharge.
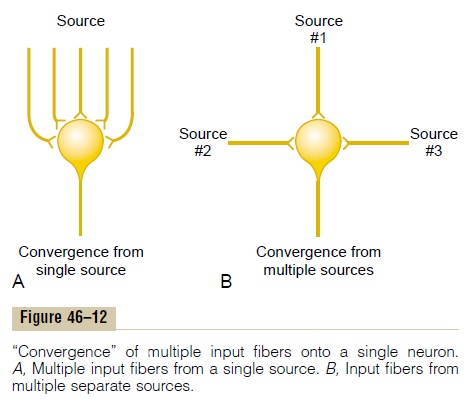
Convergence can also result from input signals (exci-tatory or inhibitory) from multiple sources, as shown in Figure 46–12B. For instance, the interneurons of the spinal cord receive converging signals from (1) periph-eral nerve fibers entering the cord, (2) propriospinal fibers passing from one segment of the cord to another, (3) corticospinal fibers from the cerebral cortex, and (4) several other long pathways descend-ing from the brain into the spinal cord.Then the signals from the interneurons converge on the anterior motor neurons to control muscle function.
Such convergence allows summation of information from different sources, and the resulting response is a summated effect of all the different types of informa-tion. Convergence is one of the important means by which the central nervous system correlates, sum-mates, and sorts different types of information.
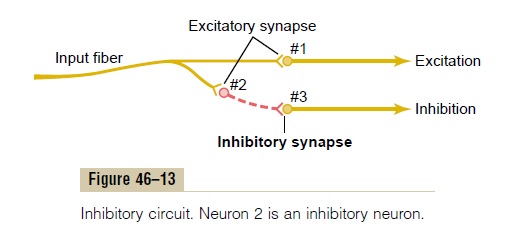
Neuronal Circuit with Both Excitatory and Inhibitory Output Signals Sometimes an incoming signal to a neuronal pool causes an output excitatory signal going in one direc-tion and at the same time an inhibitory signal going elsewhere. For instance, at the same time that an exci-tatory signal is transmitted by one set of neurons in the spinal cord to cause forward movement of a leg, an inhibitory signal is transmitted through a separate set of neurons to inhibit the muscles on the back of the leg so that they will not oppose the forward move-ment. This type of circuit is characteristic for control-ling all antagonistic pairs of muscles, and it is called the reciprocal inhibition circuit.
Figure 46–13 shows the means by which the inhibi-tion is achieved. The input fiber directly excites the excitatory output pathway, but it stimulates an inter-mediateinhibitory neuron (neuron 2), which secretes a different type of transmitter substance to inhibit the second output pathway from the pool. This type of circuit is also important in preventing overactivity in many parts of the brain.
Prolongation of a Signal by a Neuronal Pool—“Afterdischarge”
Thus far, we have considered signals that are merely relayed through neuronal pools. However, in many instances, a signal entering a pool causes a prolonged output discharge, called afterdischarge, lasting a few milliseconds to as long as many minutes after the incoming signal is over. The most important mechanisms by which afterdischarge occurs are the following.
Synaptic Afterdischarge. When excitatory synapses dis-charge on the surfaces of dendrites or soma of a neuron, a postsynaptic electrical potential develops in the neuron and lasts for many milliseconds, especially when some of the long-acting synaptic transmitter sub-stances are involved. As long as this potential lasts, it can continue to excite the neuron, causing it to trans-mit a continuous train of output impulses. Thus, as a result of this synap-tic “afterdischarge” mechanism alone, it is possible for a single instantaneous input signal to cause a sustained signal output (a series of repetitive discharges) lasting for many milliseconds.
Reverberatory (Oscillatory) Circuit as a Cause of Signal Prolon- gation. One of the most important of all circuits in theentire nervous system is the reverberatory, or oscilla-tory, circuit. Such circuits are caused by positive feed-back within the neuronal circuit that feeds back to re-excite the input of the same circuit. Consequently, once stimulated, the circuit may discharge repetitively for a long time.
Several possible varieties of reverberatory circuits are shown in Figure 46–14. The simplest, shown in Figure 46–14A, involves only a single neuron. In this case, the output neuron simply sends a collateral nerve fiber back to its own dendrites or soma to restimulate itself. Although this type of circuit probably is not an important one, theoretically, once the neuron dis-charges, the feedback stimuli could keep the neuron discharging for a protracted time thereafter.
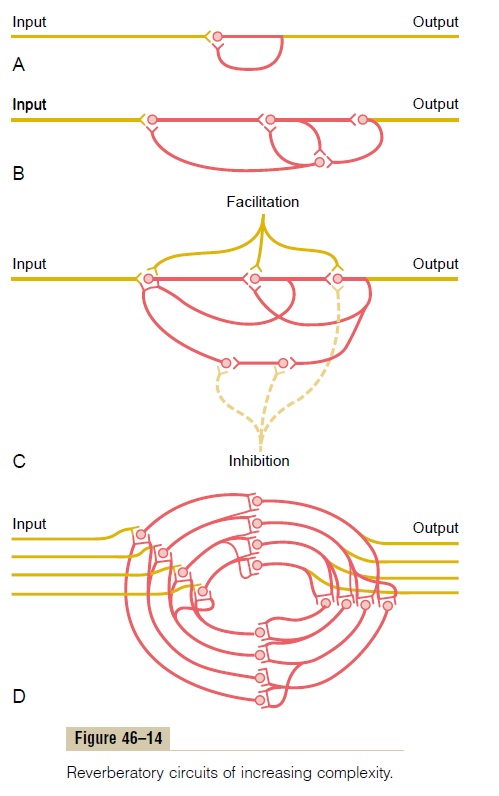
Figure 46–14B shows a few additional neurons in the feedback circuit, which causes a longer delay between initial discharge and the feedback signal. Figure 46–14Cshows a still more complex system in which both facilitatory and inhibitory fibers impinge on the reverberating circuit.A facilitatory signal enhances the intensity and frequency of reverberation, whereas an inhibitory signal depresses or stops the reverberation.
Figure 46–14D shows that most reverberating path-ways are constituted of many parallel fibers. At each cell station, the terminal fibrils spread widely. In such a system, the total reverberating signal can be either weak or strong, depending on how many parallel nerve fibers are momentarily involved in the reverberation.
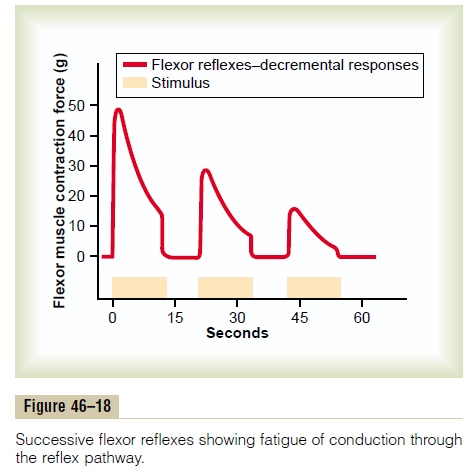
Characteristics of Signal Prolongation from a Rever- beratory Circuit. Figure 46–15 shows output signalsfrom a typical reverberatory circuit. The input stimu-lus may last only 1 millisecond or so, and yet the output can last for many milliseconds or even minutes. The figure demonstrates that the intensity of the output signal usually increases to a high value early in rever-beration and then decreases to a critical point, at which it suddenly ceases entirely. The cause of this sudden cessation of reverberation is fatigue of synaptic junc-tions in the circuit. Fatigue beyond a certain critical level lowers the stimulation of the next neuron in the circuit below threshold level so that the circuit feed-back is suddenly broken.
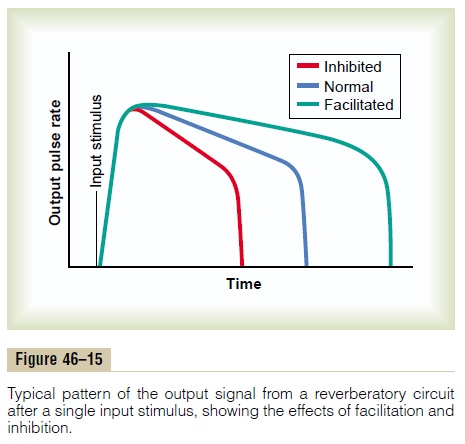
The duration of the total signal before cessation can also be controlled by signals from other parts of the brain that inhibit or facilitate the circuit. Almost these exact patterns of output signals are recorded from the motor nerves exciting a muscle involved in a flexor reflex after pain stimulation of the foot (as shown later in Figure 46–18).
Continuous Signal Output from Some Neuronal Circuits Some neuronal circuits emit output signals continu-ously, even without excitatory input signals. At least two mechanisms can cause this effect: (1) continuous intrinsic neuronal discharge and (2) continuous rever-beratory signals.
Continuous Discharge Caused by Intrinsic Neuronal Excitabil- ity. Neurons, like other excitable tissues, dischargerepetitively if their level of excitatory membrane potential rises above a certain threshold level. The membrane potentials of many neurons even normally are high enough to cause them to emit impulses con-tinually. This occurs especially in many of the neurons of the cerebellum, as well as in most of the interneu-rons of the spinal cord. The rates at which these cells emit impulses can be increased by excitatory signals or decreased by inhibitory signals; inhibitory signals often can decrease the rate of firing to zero.
Continuous Signals Emitted from Reverberating Circuits as a Means for Transmitting Information. A reverberatingcircuit that does not fatigue enough to stop reverber-ation is a source of continuous impulses. And excita-tory impulses entering the reverberating pool can increase the output signal, whereas inhibition can decrease or even extinguish the signal.
Figure 46–16 shows a continuous output signal from a pool of neurons. The pool may be emitting impulses because of intrinsic neuronal excitability or as a result
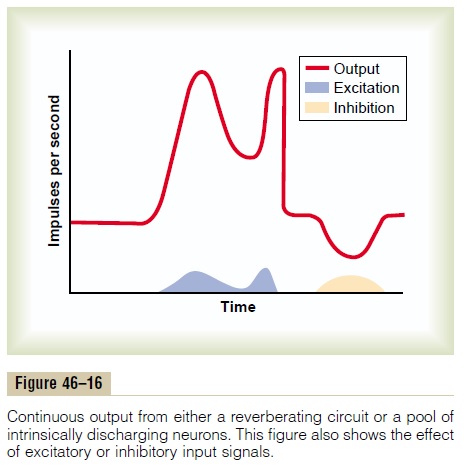
of reverberation. Note that an excitatory input signal greatly increases the output signal, whereas an inhibitory input signal greatly decreases the output. Those students who are familiar with radio trans-mitters will recognize this to be a carrier wave type of information transmission. That is, the excitatory and inhibitory control signals are not the cause of the output signal, but they do control its changing level of intensity. Note that this carrier wave system allows a decrease in signal intensity as well as an increase,whereas up to this point, the types of information transmission we have discussed have been mainly pos-itive information rather than negative information. This type of information transmission is used by the autonomic nervous system to control such functions as vascular tone, gut tone, degree of constriction of the iris in the eye, and heart rate. That is, the nerve exci-tatory signal to each of these can be either increased or decreased by accessory input signals into the rever-berating neuronal pathway.
Rhythmical Signal Output
Many neuronal circuits emit rhythmical output signals—for instance, a rhythmical respiratory signal originates in the respiratory centers of the medulla and pons. This respiratory rhythmical signal continues throughout life. Other rhythmical signals, such as those that cause scratching movements by the hind leg of a dog or the walking movements of any animal, require input stimuli into the respective circuits to initiate the rhythmical signals.
All or almost all rhythmical signals that have been studied experimentally have been found to result from reverberating circuits or a succession of sequential reverberating circuits that feed excitatory or inhibitory signals in a circular pathway from one neuronal pool to the next.
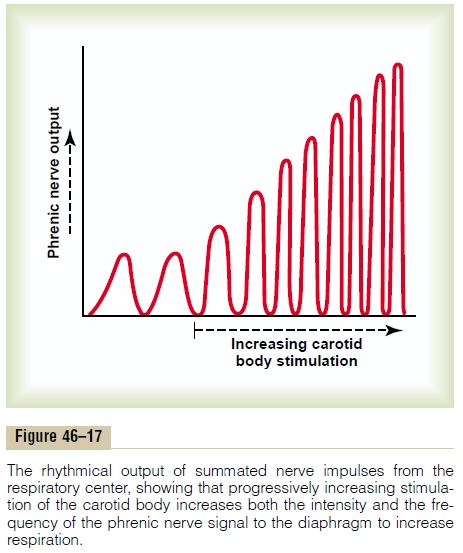
Excitatory or inhibitory signals can also increase or decrease the amplitude of the rhythmical signal output. Figure 46–17, for instance, shows changes in the respiratory signal output in the phrenic nerve. When the carotid body is stimulated by arterial oxygen deficiency, both the frequency and the amplitude of the respiratory rhythmical output signal increase progressively.
Related Topics