Chapter: Medical Physiology: Sensory Receptors, Neuronal Circuits for Processing Information
Transduction of Sensory Stimuli into Nerve Impulses
Transduction of Sensory Stimuli into Nerve Impulses
Local Electrical Currents at Nerve Endings—Receptor Potentials
All sensory receptors have one feature in common. Whatever the type of stimulus that excites the recep-tor, its immediate effect is to change the membrane electrical potential of the receptor. This change in potential is called a receptor potential.
Mechanisms of Receptor Potentials. Different receptorscan be excited in one of several ways to cause recep-tor potentials: (1) by mechanical deformation of the receptor, which stretches the receptor membrane and opens ion channels; (2) by application of a chemical to the membrane, which also opens ion channels; (3) by change of the temperature of the membrane, which alters the permeability of the membrane; or (4) by the effects of electromagnetic radiation, such as light on a retinal visual receptor, which either directly or indi-rectly changes the receptor membrane characteristics and allows ions to flow through membrane channels. It will be recognized that these four means of exciting receptors correspond in general with the different types of known sensory receptors. In all instances, the basic cause of the change in membrane potential is a change in membrane permeability of the receptor, which allows ions to diffuse more or less readily through the membrane and thereby to change the transmembrane potential.
Maximum Receptor Potential Amplitude. The maximumamplitude of most sensory receptor potentials is about 100 millivolts, but this level occurs only at an extremely high intensity of sensory stimulus. This is about the same maximum voltage recorded in action potentials and is also the change in voltage when the membrane becomes maximally permeable to sodium ions.
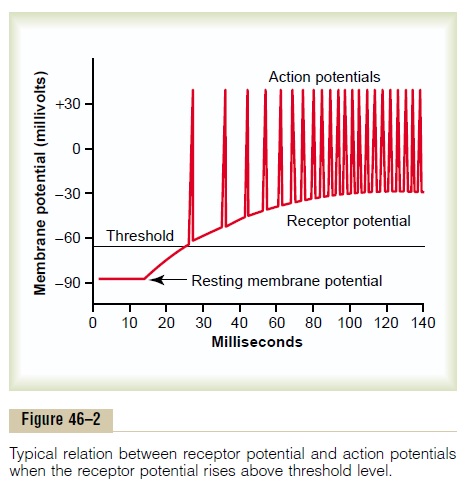
Relation of the Receptor Potential to Action Potentials. Whenthe receptor potential rises above the threshold for eliciting action potentials in the nerve fiber attached to the receptor, then action potentials occur, as illus-trated in Figure 46–2. Note also that the more the receptor potential rises above the threshold level, the greater becomes the action potential frequency.
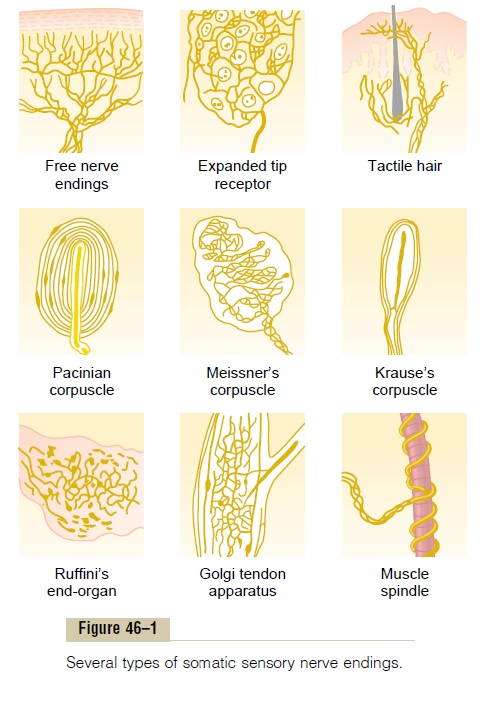
Receptor Potential of the Pacinian Corpuscle— An Example of Receptor Function The student should at this point restudy the anatomi-cal structure of the pacinian corpuscle shown in Figure 46–1. Note that the corpuscle has a central nerve fiber extending through its core. Surrounding this are mul-tiple concentric capsule layers, so that compression anywhere on the outside of the corpuscle will elongate, indent, or otherwise deform the central fiber.
Now study Figure 46–3, which shows only the central fiber of the pacinian corpuscle after all capsule layers but one have been removed. The tip of the central fiber inside the capsule is unmyelinated, but the fiber does become myelinated (the blue sheath shown in the figure) shortly before leaving the cor-puscle to enter a peripheral sensory nerve.
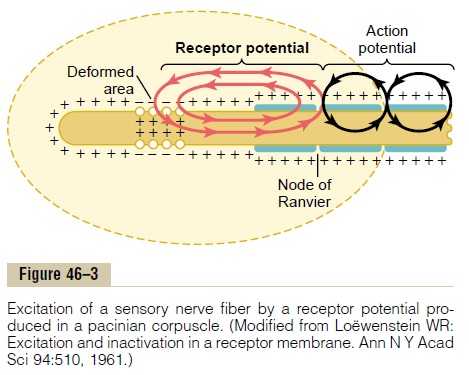
The figure also shows the mechanism by which a receptor potential is produced in the pacinian corpus-cle. Observe the small area of the terminal fiber that has been deformed by compression of the corpuscle, and note that ion channels have opened in the mem-brane, allowing positively charged sodium ions to diffuse to the interior of the fiber. This creates increased positivity inside the fiber, which is the “receptor potential.” The receptor potential in turn induces a local circuit of current flow, shown by the arrows, that spreads along the nerve fiber. At the first node of Ranvier, which itself lies inside the capsule of the pacinian corpuscle, the local current flow depolar-izes the fiber membrane at this node, which then sets off typical action potentials that are transmitted along the nerve fiber toward the central nervous system.
Relation Between Stimulus Intensity and the Receptor Poten- tial. Figure 46–4 shows the changing amplitude of thereceptor potential caused by progressively stronger mechanical compression (increasing “stimulus strength”) applied experimentally to the central core of a pacinian corpuscle. Note that the amplitude increases rapidly at first but then progressively less rapidly at high stimulus strength.
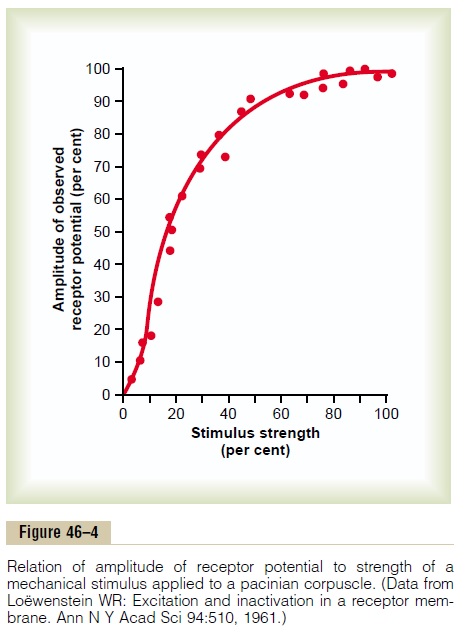
In turn, the frequency of repetitive action potentials transmitted from sensory receptors increases approx-imately in proportion to the increase in receptor potential. Putting this principle together with the data in Figure 46–4, one can see that very intense stimula-tion of the receptor causes progressively less and less additional increase in numbers of action potentials. This is an exceedingly important principle that is appli-cable to almost all sensory receptors. It allows the receptor to be sensitive to very weak sensory experi-ence and yet not reach a maximum firing rate until the sensory experience is extreme. This allows the recep-tor to have an extreme range of response, from very weak to very intense.
Adaptation of Receptors
Another characteristic of all sensory receptors is that they adapt either partially or completely to any con-stant stimulus after a period of time. That is, when a continuous sensory stimulus is applied, the receptor responds at a high impulse rate at first and then at a progressively slower rate until finally the rate of action potentials decreases to very few or often to none at all.
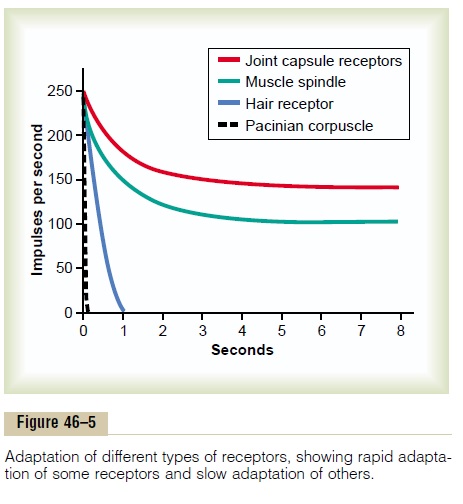
Figure 46–5 shows typical adaptation of certain types of receptors. Note that the pacinian corpuscle adapts extremely rapidly and hair receptors adapt within a second or so, whereas some joint capsule and muscle spindle receptors adapt slowly.
Furthermore, some sensory receptors adapt to a far greater extent than others. For example, the pacinian corpuscles adapt to “extinction” within a few hun-dredths of a second, and the receptors at the bases of the hairs adapt to extinction within a second or more. It is probable that all other mechanoreceptors eventu-ally adapt almost completely, but some require hours or days to do so, for which reason they are called “non-adapting” receptors. The longest measured time for complete adaptation of a mechanoreceptor is about 2 days, which is the adaptation time for many caro-tid and aortic baroreceptors. Conversely, some of the nonmechanoreceptors—the chemoreceptors and pain receptors, for instance—probably never adapt completely.
Mechanisms by Which Receptors Adapt. The mechanism ofreceptor adaptation is different for each type of recep-tor, in much the same way that development of a receptor potential is an individual property. For instance, in the eye, the rods and cones adapt by chang-ing the concentrations of their light-sensitive chemi-cals.
In the case of the mechanoreceptors, the receptor that has been studied in greatest detail is the pacinian corpuscle. Adaptation occurs in this receptor in two ways. First, the pacinian corpuscle is a viscoelastic structure so that when a distorting force is suddenly applied to one side of the corpuscle, this force is instantly transmitted by the viscous component of the corpuscle directly to the same side of the central nerve fiber, thus eliciting a receptor potential. However, within a few hundredths of a second, the fluid within the corpuscle redistributes, so that the receptor poten-tial is no longer elicited. Thus, the receptor potential appears at the onset of compression but disappears within a small fraction of a second even though the compression continues.
The second mechanism of adaptation of the pacin-ian corpuscle, but a much slower one, results from a process called accommodation, which occurs in the nerve fiber itself. That is, even if by chance the central core fiber should continue to be distorted, the tip of the nerve fiber itself gradually becomes “accommo-dated” to the stimulus. This probably results from progressive “inactivation” of the sodium channels in the nerve fiber membrane, which means that sodium current flow through the channels causes them gradu-ally to close, an effect that seems to occur for all or most cell membrane sodium channels.
Presumably, these same two general mechanisms of adaptation apply also to the other types of mechano-receptors. That is, part of the adaptation results from readjustments in the structure of the receptor itself, and part from an electrical type of accommodation in the terminal nerve fibril.
Slowly Adapting Receptors Detect Continuous Stimulus Strength—The “Tonic” Receptors. Slowly adapting recep-tors continue to transmit impulses to the brain as long as the stimulus is present (or at least for many minutes or hours). Therefore, they keep the brain constantly apprised of the status of the body and its relation to its surroundings. For instance, impulses from the muscle spindles and Golgi tendon apparatuses allow the nervous system to know the status of muscle con-traction and load on the muscle tendon at each instant.
Other slowly adapting receptors include (1) recep-tors of the macula in the vestibular apparatus, (2) pain receptors, (3) baroreceptors of the arterial tree, and (4) chemoreceptors of the carotid and aortic bodies.
Because the slowly adapting receptors can continue to transmit information for many hours, they are called tonic receptors.
Rapidly Adapting Receptors Detect Change in Stimulus Strength—The “Rate Receptors,” “Movement Receptors,” or “Phasic Receptors.” Receptors that adapt rapidlycannot be used to transmit a continuous signal because these receptors are stimulated only when the stimulus strength changes. Yet they react strongly while achange is actually taking place. Therefore, these recep-tors are called rate receptors, movement receptors, or phasic receptors. Thus, in the case of the pacinian cor-puscle, sudden pressure applied to the tissue excites this receptor for a few milliseconds, and then its exci-tation is over even though the pressure continues. But later, it transmits a signal again when the pressure is released. In other words, the pacinian corpuscle is exceedingly important in apprising the nervous system of rapid tissue deformations, but it is useless for trans-mitting information about constant conditions in the body.
Importance of the Rate Receptors—Their Predictive Function. If one knows the rate at which some change in bodily status is taking place, one can predict in one’s mind the state of the body a few seconds or even a few minutes later. For instance, the receptors of the semicircular canals in the vestibular apparatus of the ear detect the rate at which the head begins to turn when one runs around a curve. Using this information, a person can predict how much he or she will turn within the next 2 seconds and can adjust the motion of the legs aheadof time to keep from losing balance. Likewise, recep-tors located in or near the joints help detect the rates of movement of the different parts of the body. For instance, when one is running, information from the joint rate receptors allows the nervous system to predict where the feet will be during any precise frac-tion of the next second. Therefore, appropriate motor signals can be transmitted to the muscles of the legs to make any necessary anticipatory corrections in posi-tion so that the person will not fall. Loss of this pre-dictive function makes it impossible for the person to run.
Related Topics