Chapter: Medical Physiology: Resistance of the Body to Infection: II. Immunity and Allergy
Specific Attributes of the B-Lymphocyte System - Humoral Immunity and the Antibodies
Specific Attributes of the B-Lymphocyte System - Humoral Immunity and the Antibodies
Formation of Antibodies by Plasma Cells. Before exposureto a specific antigen, the clones of B lymphocytes remain dormant in the lymphoid tissue. On entry of a foreign antigen, macrophages in the lymphoid tissue phagocytize the antigen and then present it to adjacent B lymphocytes. In addition, the antigen is presented to T cells at the same time, and activated helper T cells are formed. These helper cells also contribute to extreme activation of the B lymphocytes, as we discuss more fully later.
Those B lymphocytes specific for the antigen imme-diately enlarge and take on the appearance of lymphoblasts. Some of the lymphoblasts further dif-ferentiate to form plasmablasts, which are precursors of plasma cells. In the plasmablasts, the cytoplasm expands and the rough endoplasmic reticulum vastly proliferates. The plasmablasts then begin to divide at a rate of about once every 10 hours for about nine divi-sions, giving in 4 days a total population of about 500 cells for each original plasmablast. The mature plasma cell then produces gamma globulin antibodies at an extremely rapid rate—about 2000 molecules per second for each plasma cell. In turn, the antibodies are secreted into the lymph and carried to the circulating blood.This process continues for several days or weeks until finally exhaustion and death of the plasma cells occur.
Formation of “Memory” Cells—Difference Between Primary Response and Secondary Response. A few of the lym-phoblasts formed by activation of a clone of B lym-phocytes do not go on to form plasma cells but instead form moderate numbers of new B lymphocytes similar to those of the original clone. In other words, the B-cell population of the specifically activated clone becomes greatly enhanced, and the new B lympho-cytes are added to the original lymphocytes of the same clone. They also circulate throughout the body to populate all the lymphoid tissue; immunologically, however, they remain dormant until activated once again by a new quantity of the same antigen. These lymphocytes are called memory cells. Subsequent exposure to the same antigen will cause a much more rapid and much more potent antibody response this second time around, because there are many more memory cells than there were original B lymphocytes of the specific clone.
Figure 34–2 shows the differences between the primary response for forming antibodies that occurs onfirst exposure to a specific antigen and the secondaryresponse that occurs after second exposure to the sameantigen. Note the 1-week delay in the appearance of the primary response, its weak potency, and its short life. The secondary response, by contrast, begins rapidly after exposure to the antigen (often within hours), is far more potent, and forms antibodies for many months rather than for only a few weeks. The
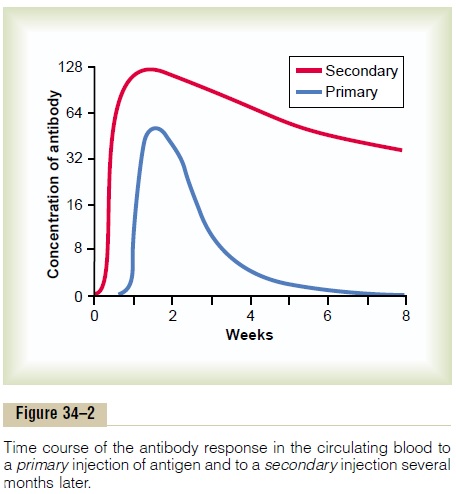
increased potency and duration of the secondary response explain why immunization is usually accom-plished by injecting antigen in multiple doses with periods of several weeks or several months between injections.
Nature of the Antibodies
The antibodies are gamma globulins called immuno-globulins (abbreviated as Ig), and they have molecu-lar weights between 160,000 and 970,000. They usually constitute about 20 per cent of all the plasma proteins.
All the immunoglobulins are composed of combi-nations of light and heavy polypeptide chains. Most are a combination of two light and two heavy chains, as shown in Figure 34–3. However, some of the immunoglobulins have combinations of as many as 10 heavy and 10 light chains, which gives rise to high-molecular-weight immunoglobulins. Yet, in all immunoglobulins, each heavy chain is paralleled by a light chain at one of its ends, thus forming a heavy-light pair, and there are always at least 2 and as many as 10 such pairs in each immunoglobulin molecule.
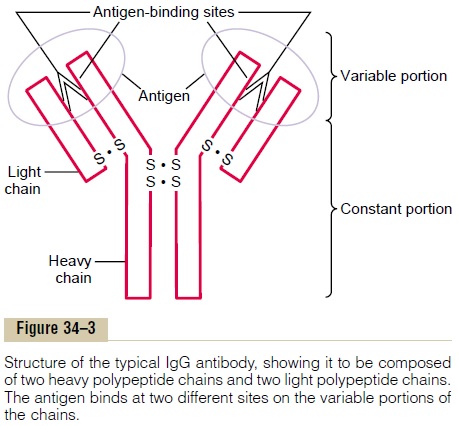
Figure 34–3 shows by the circled area a designated end of each light and heavy chain, called the variableportion; the remainder of each chain is called the con-stant portion. The variable portion is different for eachspecificity of antibody, and it is this portion that attaches specifically to a particular type of antigen.The constant portion of the antibody determines other properties of the antibody, establishing such factors as diffusivity of the antibody in the tissues, adherence of the antibody to specific structures within the tissues, attachment to the complement complex, the ease with which the antibodies pass through membranes, and other biological properties of the antibody.
Specificity of Antibodies. Each antibody is specific for aparticular antigen; this is caused by its unique struc-tural organization of amino acids in the variable por-tions of both the light and heavy chains. The amino acid organization has a different steric shape for each antigen specificity, so that when an antigen comes in contact with it, multiple prosthetic groups of the antigen fit as a mirror image with those of the anti-body, thus allowing rapid and tight bonding between the antibody and the antigen. When the antibody is highly specific, there are so many bonding sites that the antibody-antigen coupling is exceedingly strong, held together by (1) hydrophobic bonding, (2) hydrogen bonding, (3) ionic attractions, and (4) van der Waals forces. It also obeys the thermodynamic mass action law.

Kais called the affinity constant and is a measure ofhow tightly the antibody binds with the antigen.
Note, especially, in Figure 34–3 that there are two variable sites on the illustrated antibody for attach-ment of antigens, making this type of antibody biva-lent. A small proportion of the antibodies, whichconsist of combinations of up to 10 light and 10 heavy chains, have as many as 10 binding sites.
Classes of Antibodies. There are five general classes ofantibodies, respectively named IgM, IgG, IgA, IgD, and IgE. Ig stands for immunoglobulin, and the other five respective letters designate the respective classes.
For the purpose of our present limited discussion, two of these classes of antibodies are of particular importance: IgG, which is a bivalent antibody and con-stitutes about 75 per cent of the antibodies of the normal person, and IgE, which constitutes only a small percentage of the antibodies but is especially involved in allergy. The IgM class is also interesting because a large share of the antibodies formed during the primary response are of this type.
These antibodies have 10 binding sites that make them exceedingly effective in protecting the body against invaders, even though there are not many IgM antibodies.
Mechanisms of Action of Antibodies
Antibodies act mainly in two ways to protect the body against invading agents: (1) by direct attack on the invader and (2) by activation of the “complement system” that then has multiple means of its own for destroying the invader.
Direct Action of Antibodies on Invading Agents. Figure 34–4shows antibodies (designated by the red Y-shaped bars) reacting with antigens (designated by the shaded objects). Because of the bivalent nature of the anti-bodies and the multiple antigen sites on most invad-ing agents, the antibodies can inactivate the invading agent in one of several ways, as follows:
1. Agglutination, in which multiple large particleswith antigens on their surfaces, such as bacteria or red cells, are bound together into a clump
2. Precipitation, in which the molecular complex ofsoluble antigen (such as tetanus toxin) and antibody becomes so large that it is rendered insoluble and precipitates
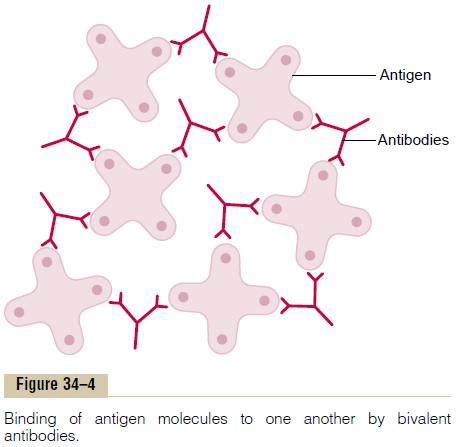
3. Neutralization, in which the antibodies cover thetoxic sites of the antigenic agent
4. Lysis, in which some potent antibodies areoccasionally capable of directly attacking membranes of cellular agents and thereby cause rupture of the agent
These direct actions of antibodies attacking the anti-genic invaders often are not strong enough to play a major role in protecting the body against the invader.
Most of the protection comes through the amplifying effects of the complement system described next.
Complement System for Antibody Action “Complement” is a collective term that describes a system of about 20 proteins, many of which are enzyme precursors. The principal actors in this system are 11 proteins designated C1 through C9, B, and D, shown in Figure 34–5. All these are present normally among the plasma proteins in the blood as well as among the proteins that leak out of the capillaries into the tissue spaces. The enzyme precursors are normally inactive, but they can be activated mainly by the so-called classic pathway.
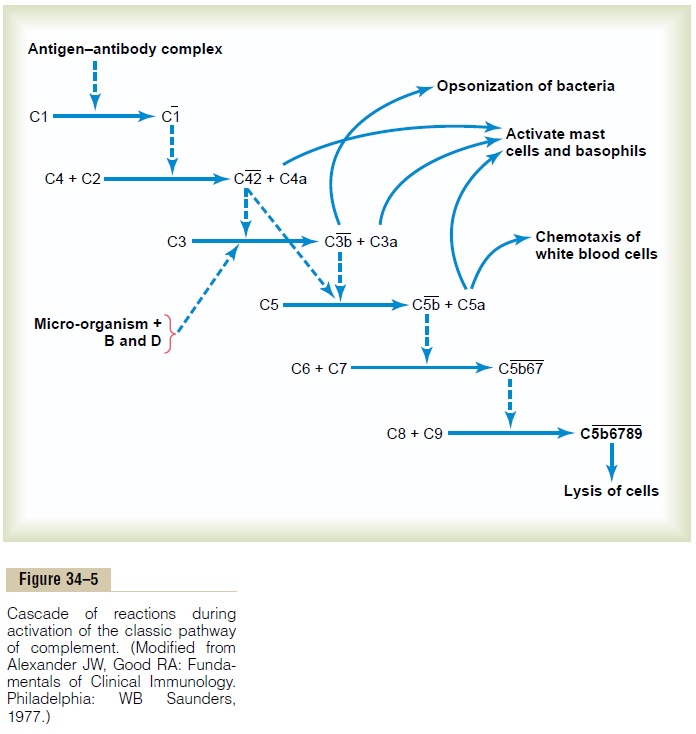
Classic Pathway. The classic pathway is initiated by anantigen-antibody reaction. That is, when an antibody binds with an antigen, a specific reactive site on the “constant” portion of the antibody becomes uncov-ered, or “activated,” and this in turn binds directly with the C1 molecule of the complement system, setting into motion a “cascade” of sequential reactions, shown in Figure 34–5, beginning with activation of the proen-zyme C1 itself. The C1 enzymes that are formed then activate successively increasing quantities of enzymes inthe later stages of the system, so that from a small beginning, an extremely large “amplified” reaction occurs. Multiple end products are formed, as shown to the right in the figure, and several of these cause important effects that help to prevent damage to the body’s tissues caused by the invading organism or toxin. Among the more important effects are the following:
1. Opsonization and phagocytosis. One of theproducts of the complement cascade, C3b, strongly activates phagocytosis by both neutrophils and macrophages, causing these cells to engulf the bacteria to which the antigen-antibody complexes are attached. This process is called opsonization. It often enhances the number of bacteria that can be destroyed by many hundredfold.
2. Lysis. One of the most important of all theproducts of the complement cascade is the lyticcomplex, which is a combination of multiplecomplement factors and designated C5b6789. This has a direct effect of rupturing the cell membranes of bacteria or other invading organisms.
3. Agglutination. The complement products alsochange the surfaces of the invading organisms, causing them to adhere to one another, thus promoting agglutination.
4. Neutralization of viruses. The complementenzymes and other complement products can attack the structures of some viruses and thereby render them nonvirulent.
5. Chemotaxis. Fragment C5a initiates chemotaxis ofneutrophils and macrophages, thus causing large numbers of these phagocytes to migrate into the tissue area adjacent to the antigenic agent.
6. Activation of mast cells and basophils. FragmentsC3a, C4a, and C5a activate mast cells and basophils, causing them to release histamine, heparin, and several other substances into the local fluids. These substances in turn cause increased local blood flow, increased leakage of fluid and plasma protein into the tissue, and other local tissue reactions that help inactivate or immobilize the antigenic agent. The same factors play a major role in inflammation and in allergy, as we discuss later.
7. Inflammatory effects. In addition to inflammatoryeffects caused by activation of the mast cells and basophils, several other complement products contribute to local inflammation. These products cause (1) the already increased blood flow to increase still further, (2) the capillary leakage of proteins to be increased, and (3) the interstitial fluid proteins to coagulate in the tissue spaces, thus preventing movement of the invading organism through the tissues.
Related Topics