Chapter: Essential Microbiology: Procaryote Diversity
Phylum: Proteobacteria
Domain: Bacteria
This is divided into 23 phyla (Table 7.2), the more important of which are discussed in the following pages, according to their description in the second edition of Bergey. As with the Archaea, many other forms are known only through molecular analysis and it is estimated that these represent at least another 20 phyla.

Phylum: Proteobacteria
We start our survey of the Bacteria with the Proteobacteria. This is by far the biggest single phylum. The size of the group is matched by its diversity, both morphological and physiologi-cal; most forms of metabolism are represented, and the wide range of morphological forms gives rise to the groupŌĆÖs name. (Proteus was a mythological Greek god who was able to assume many different forms.) The reason such a diverse range of organisms
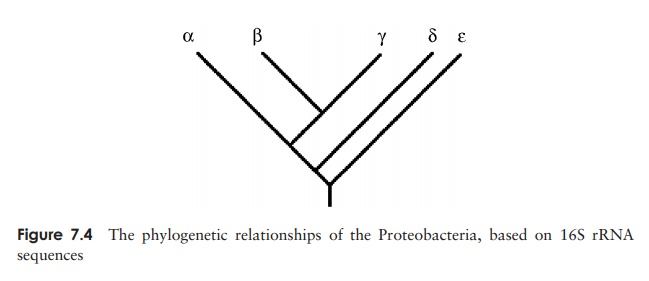
At the time of writing more than 460 genera and 1600 species had been iden-tified, all of them Gram-negative and representing almost half of all accepted bacterial genera. These include many of the best known Gram-negative bacteria of medical, in-dustrial and agricultural importance. For taxonomic purposes, the Proteobacteria have been divided into five classes reflecting their presumed lines of descent and termed the Alphaproteobacteria, Betaproteobacteria, Gammaproteobacteria, Deltaproteobacteria and Epsilonproteobacteria (Figure 7.4). It should be stressed that because classification is based on molecular relatedness rather than shared phenotypic traits, few if any mor-phological or physiological properties can be said to be characteristic of all members of each class. Equally, organisms united by a particular feature may be found in more than one of the proteobacterial classes, for example nitrifying bacteria are to be found in the ╬▒, ╬▓ and ╬│ Protobacteria. For this reason, in the following paragraphs we describe theProteobacteria in terms of their phenotypic characteristics rather than attempt to group them phylogenetically.
Photosynthetic Proteobacteria
The purple sulphur and purple non-sulphur bacteria are the only members of the Pro-teobacteria to have retained the photosynthetic ability of their presumed ancestor. The type of photosynthesis they carry out, however, is quite distinct from that carried out by plants, algae and cyanobacteria, differing in two important respects:
┬Ę it is anoxygenic ŌĆō no oxygen is produced by the process
┬Ę it utilizes bacteriochlorophyll a and/or b, which have different absorbance prop-erties to chlorophylls a and b.
Like organisms that carry out green photosynthesis, however, they incorporate CO2 into carbohydrate by means of the Calvin cycle .
All are at least facultatively anaerobic, and are typically found in sediments of stagnant lakes and salt marsh pools, where they may form extensive coloured blooms. Because the absorption spectrum of bacteriochlorophylls lies mostly in the infrared part of the spectrum, they are able to utilise light energy that penetrates beyond the surface layers of water.
The coloration, ranging from orange/brown to purple, is due to the presence of carotenoid pigments such as lycopene and spirillixanthin, which mask the blue/green colour of the bacteriochlorophylls. The photosynthetic pigments are located on highly folded extensions of the plasma membrane. Photosynthetic proteobacteria include rods, cocci and spiral forms.
Under anaerobic conditions, the purple sulphur bacteria typically utilise hydrogen sulphide or elemental sulphur as an electron donor for the reduction of CO2.
H2S +CO2ŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌåÆ (CH2O)n+ S0
S0+CO2+ H2O ŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌåÆ (CH2O)n+ H2SO4
Many store sulphur in the form of intracellular granules. The purple sulphur bacteria all belong to the ╬│ -Protobacteria. They are typically found in surface muds, and sulphur springs, habitats that provide the right combination of light and anaerobic conditions.
Representative genera: Thiospirillum, Chromatium
The purple non-sulphur bacteria were distinguished from the above group because of their apparent inability to use H2S as an electron donor. It is now known, however, that the majority can do this, but are able to tolerate much lower concentrations in comparison with the purple sulphur bacteria. The purple non-sulphur bacteria are fac-ultative anaerobes able to grow as photoheterotrophs, that is, with light as an energy source and a range of organic molecules such as carbohydrates and organic acids as sources of both carbon and electrons. In addition, many are able to grow aerobically as chemoheterotrophs in the absence of light. Under present classification systems, purple nonsulphur bacteria are divided between the ╬▒- and ╬▓-Proteobacteria.
Representative genera: Rhodospirillum, Rhodopseudomonas
Nitrifying Proteobacteria
This group comprises aerobic Gram-negative chemolithoautotrophs that derive their energy from the oxidation of inorganic nitrogen compounds (either ammonia or nitrite), and their carbon from CO2. The nitrifying bacteria are represented in both the ╬▒- and ╬▓-Proteobacteria.
The oxidation of ammonia through to nitrate is a two-stage process, with specific bacteria carrying out each stage (ammonia to nitrite and nitrite to nitrate). This is reflected in the generic names of the bacteria, bearing the prefix Nitroso- or Nitro-according to whether they carry out the first or second reaction. Nitrifying bacteria play an essential role in the cycling of nitrogen in terrestrial, marine and freshwater habitats. Nitrite, which is toxic to many forms of life, rarely accumulates in the environment, due to the activity of the Nitrobacteria. As was the case with the purple photosynthetic bacteria, several cell forms are represented among the nitrifiers.
Representative genera: Nitrosomonas (NH4+ ŌłÆŌłÆŌłÆŌłÆŌłÆŌåÆ NO2ŌłÆ)
Nitrobacter (NO2ŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌåÆNO3ŌłÆ)
Iron- and sulphur-oxidising Proteobacteria
Two further groups of environmentally significant chemolithoautotrophs derive their energy through the oxidation of reduced iron and sulphur respectively.
Among the sulphur oxidisers, perhaps the best studied are members of the genus Acidithiobacillus*, which includes extreme acidophiles such as A. thiooxidans that arecapable of growth at a pH as low as 1.0! These may utilise sulphur in its elemental form, as H2S, metal sulphides, or other forms of reduced sulphur such as thiosulphate:
2S0+ 3O2+ 2H2O ŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌåÆ 2H2SO4
S2ŌłÆ+ 4O2ŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌåÆ 2SO42ŌłÆ
S2O32ŌłÆ+ 2O2+ H2O ŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌåÆ 2SO42ŌłÆ+ 2H+
The result of all these reactions is the production of sulphuric acid and a lowering of the environmental pH. Bacteria such as these are responsible for the phenomenon of acidmine drainage. A particularly valuable organism in the latter context is A. ferrooxidans, due to its ability to use not only reduced sulphur compounds as energy sources, but also reduced iron .
A second group of sulphur oxidisers are bacteria that exist not as single cells, but join to form filaments, the best known of which is Beggiotoa. These are typically found in sulphur springs, marine sediments and hydrothermal vents at the bottom of the sea.
Acidithiobacillus ferrooxidans is also an example of an iron oxidiser. At normalphysiological pH values and in the presence of oxygen, reduced iron (iron II, Fe2+ ) is spontaneously oxidised to the oxidised form (iron III, Fe3+ ). Under very acidic con-ditions, the iron remains in its reduced form, unless acted on by certain bacteria. A.ferrooxidans; is an obligate aerobe able to use iron II as an energy source, converting itto iron III at an optimum pH range of around 2:
2Fe2++1/2O2+ 2H+ŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌåÆ Fe3++ H2O
Gallionella ferruginea, on the other hand, grows around neutrality in oxygen-poorenvironments such as bogs and iron springs. Ferric hydroxide is excreted from the cell and deposited on a stalk-like structure projecting from, and much bigger than, the cell itself. This gives the macroscopic impression of a mass of red/brown twisted filaments.
Representative genera: Acidithiobacillus, Beggiotoa (sulphur oxidisers) , Leptospirillum, Gallionella (iron oxidisers)
Hydrogen-oxidising Proteobacteria
This diverse group of bacteria are united by their ability to derive energy by using hydrogen gas as a donor of electrons, and oxygen as an acceptor:
2H2+ O2ŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌåÆ 2H2O
Nearly all the members of this group are facultative chemolithotrophs, i.e. they can also grow as heterotrophs, utilising organic compounds instead of CO2 as their carbon source, and indeed most grow more efficiently in this way.
Representative genera: Alcaligenes, Ralstoni
Nitrogen-fixing Proteobacteria
The ╬▒-Proteobacteria includes certain genera of nitrogen-fixing bacteria. These are able to fix (reduce)atmospheric N2 as NH4+ for subsequent incorporation into cellular materials, a process that requires a consid-erable input of energy in the form of ATP:
N2 + 8eŌłÆ + 8H+ + 16ATP ŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌåÆ, 2NH4+ + 16ADP + 16Pi
Nitrogen-fixing bacteria may be free-living in the soil (e.g. Azotobacter), or form a symbiotic relationship with cells on the root hairs of leguminous plants such as peas, beans and clover (e.g. Rhizobium). The nitrogenase responsible for the reaction (actually a complex of two enzymes) is highly sensitive to oxygen; many nitrogen fixers are anaerobes, while others have devised ways of keeping the cell interior oxygen-free.
Closely related to Rhizobium, but unable to fix nitrogen, are members of the genus Agrobacterium. Like Rhizobium, these enter the tissues of plants, but instead of forminga mutually beneficial association, cause cell proliferation and tumour formation. A.tumefaciens has proved to be a valuable tool in the genetic engineering of plants.
Representative genera: Rhizobium, Azotobacter
Methanotrophic Proteobacteria
In discussing the Archaea earlier, we encountered species capable of the production of methane, a gas found widely in such diverse locations as marshes, sewage sludge and animal intestines. Certain proteobac-teria are able to utilise this methane as a carbon and energy source and are known as methanotrophs.
Methanotrophs are strict aerobes, requiring oxy-gen for the oxidation of methane. The methane-generating bacteria, however, as weŌĆÖve seen are anaer-obes; methanotrophs are consequently to be found at aerobic/anaerobic interfaces such as topsoil, where they
can find both the oxygen and the methane they require. The methane is firstly oxidised to methanol, then to formaldehyde, by means of separate enzyme systems. Some of the carbon in formaldehyde is assimilated into organic cellular material, while some is further oxidised to carbon dioxide.
Bacteria able to utilise other single-carbon com-pounds such as methanol (CH3OH) or methylamine (CH3NH2) are termed methylotrophs. Depending on whether they possess the enzyme methane monooxyge-nase (MMO), they may also be methanotrophs.
Representative genera: Methylomonas, Methylococcus
Sulphate- and sulphur-reducing Proteobacteria
Some 20 genera of anaerobic ╬┤-Proteobacteria reduce either elemental sulphur or oxi-dised forms of sulphur such as sulphate to hydrogen sulphide. Organic compounds such as pyruvate, lactate or certain fatty acids act as electron donors:
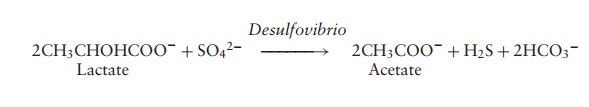
Sulphate- and sulphur-reducers are found in anaerobic muds and play an important role in the global sulphur cycle.
Representative genera: Desulfovibrio (sulphate), Desulfuromonas (sulphur)
Enteric Proteobacteria
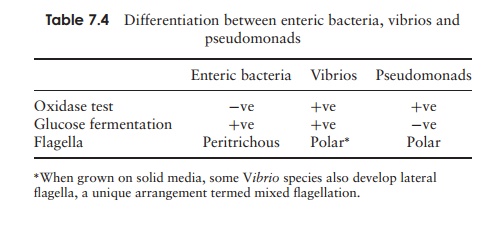
This is a large group of rod-shaped bacteria, mostly motile by means of peritrichous flagella, which all belong to the ╬│ -Proteobacteria. They are facultative aerobes, charac-terised by their ability in anaerobic conditions to carry out fermentation of glucose and other sugars to give a variety of products. The nature of these products allows division into two principal groups, the mixed acid fermenters and the butanediol fermenters (Figure 6.23). All the enteric bacteria test negative for cytochrome c oxidase (see Vibrio and related genera below). In view of their similar appearance, members of the group are distinguished from one another largely by means of their biochemical characteris-tics. An unknown isolate is subjected to a series of tests including its ability to utilise substrates such as lactose and citrate, convert tryptophan to indole, and hydrolyse urea. On the basis of its response to each test, a characteristic profile can be built up for the isolate, and matched against those of known species (see Table 7.3).


The most thoroughly studied of all bacteria, Escherichia coli (E. coli) is a member of this group, as are a number of important pathogens of humans such as Salmonella, Shigella and Yersinia (the causative agent of plague).
Representative genera: Escherichia, Enterobacter
Vibrio and related genera
A few other genera, including Vibrio and Aeromonas, are also facultative anaerobes able to carry out the fermentative reactions described above, but are differentiated from the enteric bacteria by being oxidase-positive (Table 7.4). Vibrio and Photobacteriumboth include examples of marine bioluminescentspecies; these are widely found both in seawater and associated with fish and other marine life. The lumi-nescence, which requires the presence of oxygen, is due to an oxidation reaction carried out by the enzyme luciferase.
Vibrio cholerae is the causative agent of cholera, a debilitating and often fatal formof acute diarrhoea transmitted in faecally contaminated water. It remains a major killer in many third world countries. Several species of Vibrio, including V. cholerae, have been shown to possess two circular chromosomes instead of the usual one.
Representative genera: Vibrio, Aeromonas
The Pseudomonads
Members of this group of proteobacteria, the most im-portant genus of which is Pseudomonas, are straight or curved rods with polar flagella. They are chemo-heterotrophs that generally utilise the EntnerŌĆōDoudoroff pathway rather than glycolysis for the oxidation of hexoses. They are differentiated from the enteric bacteria (Table 7.4) by being oxidase-positive and incapable of fermentation. A characteristic of many pseudomonads is the ability to utilise an extremely wide range of organic compounds (maybe over 100!) for car-bon and energy, something that makes them very im-portant in the recycling of carbon in the environment. Several species are significant pathogens of animals and plants; Pseudomonas aeruginosa is an effective coloniser of wounds and burns in humans, while P. syringae causes chlorosis (yellowing of leaves) in a range of plants. Because of their ability to grow at low temperatures, a number of pseudomonads are important in the spoilage of food.
Although most species carry out aerobic respiration with oxygen as the terminal electron acceptor, a few are capable of substituting nitrate.
Acetic acid bacteria
Acetobacter and Gluconobacter are two genera of the╬▒-Proteobacteria that convertethanol into acetic acid, a highly significant reaction in the food and drink industries. Both genera are strict aerobes, but unlike Acetobacter, which can oxidise the acetic acid right through to carbon dioxide and water, Gluconobacter lacks all the enzymes of the TCA cycle, and cannot oxidise it further.
Acetobacter species also have the ability, rare in bacteria, to synthesise cellulose;the cells become surrounded by a mass of extracellular fibrils, forming a pellicle at the surface of an unshaken liquid culture.
Representative genera: Acetobacter, Gluconobacter
Stalked and budding Proteobacteria
The members of this group of aquatic Proteobacteria differ noticeably in their appear-ance from typical bacteria by their possession of extracellular extensions known as prosthecae; these take a variety of forms but are always narrower than the cell itself.They are true extensions of the cell, containing cytoplasm, rather than completely ex-tracellular appendages.
In the stalked bacteria such as Caulobacter (Figure 7.5), the prostheca serves both as a means of attaching the cell to its substratum, and to enhance nutrient absorption by
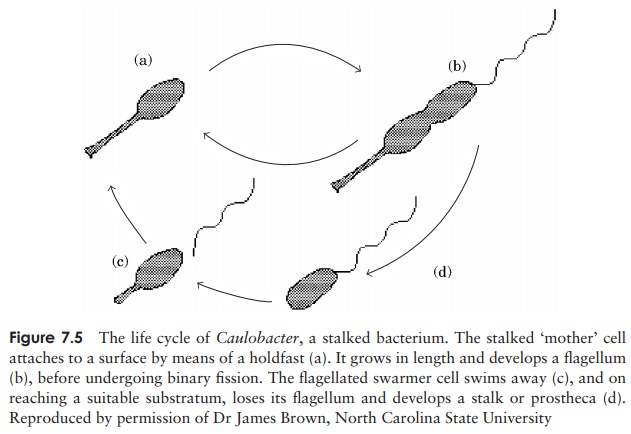
increasing the surface area-to-volume ratio of the cell. The latter enables such bacteria to live in waters containing extremely low levels of nutrient. Caulobacter lives part of its life cycle as a free-swimming swarmer cell with no prostheca but instead a flagellum for mobility.
The iron oxidiser Gallionella (see Nitrifying Proteobacteria above) may be regarded as a stalked bacterium, however it is not truly prosthecate, as its stalk does not contain cytoplasm.
In the budding bacteria, the prostheca is involved in a distinctive form of reproduc-tion, in which two cells of unequal size are produced (c.f. typical binary fission, which results in two identical daughter cells). The daughter cell buds off from the mother cell, either directly, or as Hyphomicrobium spp. at the end of a hypha (stalk) (Figure 7.6). Once detached, the daughter cell grows to full size and eventually produces its own buds. Hyphomicrobium is a methanotroph and a methylotroph, so it also belongs to the methanotrophs described earlier.

In some bacteria, more than one prostheca is found per cell; these polyprosthecate forms include the genus Stella, whose name (ŌĆśa starŌĆÖ) derives from its six symmetrically arranged buds.
Representative genera: Caulobacter, Hyphomicrobium
Sheathed Proteobacteria
Some genera of ╬▓ Proteobacteria exist as chains of cells surrounded by a tube-like sheath, made up of a carbohydrate/protein/lipid complex. In some cases, the sheath contains deposits of manganese oxide or ferric hydroxide, which may be the product of chemical or biological oxidation. Empty sheaths encrusted with oxides may remain long after the bacterial cells have died off or been released. As with the stalked bacteria the sheath helps in the absorption of nutrients, and may also offer protection against predators.
The sheathed bacteria have a relatively complex life cycle. They live in flowing water, and attach with one end of the chain to, for example, a plant or rock. Free-swimming single flagellated cells are released from the distal end and settle at another location, where a new chain and sheath are formed (Figure 7.7).
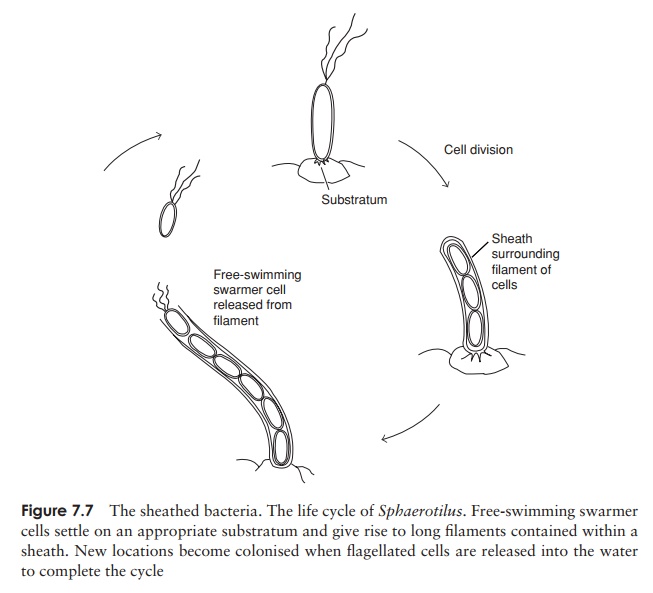
Sphaerotilus forms thick ŌĆśstreamersŌĆÖ in polluted water, and is a familiar sight aroundsewage outlets.
Representative genera: Sphaerotilus, Leptothrix
Predatory Proteobacteria
Bdellovibrio is a unique genus belonging to the╬┤-Proteobacteria. It is a very small comma-shaped bac-terium, which actually attacks and lives inside other Gram-negative bacteria (Figure 7.8). Powered by its flag-ellum, it collides with its prey at high speed and pene-trates even thick cell walls by a combination of enzyme secretion and mechanical boring. It takes up residence in the periplasmic space, between the plasma membrane and cell wall. The hostŌĆÖs nucleic acid and protein syn-thesis cease, and its macromolecules are degraded, pro-viding nutrients for the invader, which grows into a long
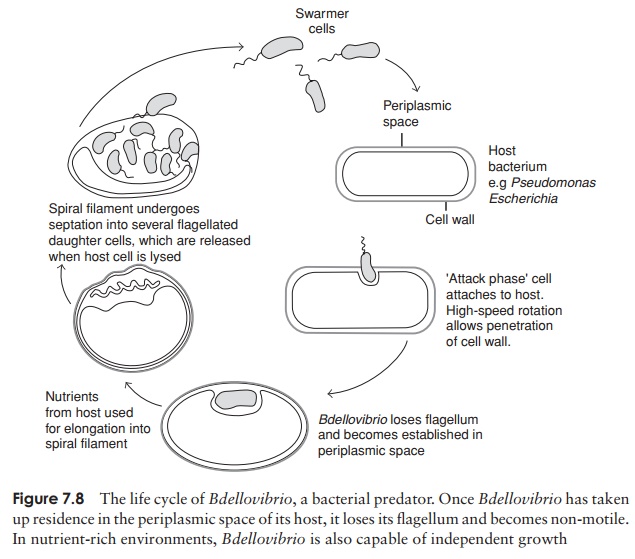
Representative genus: Bdellovibrio
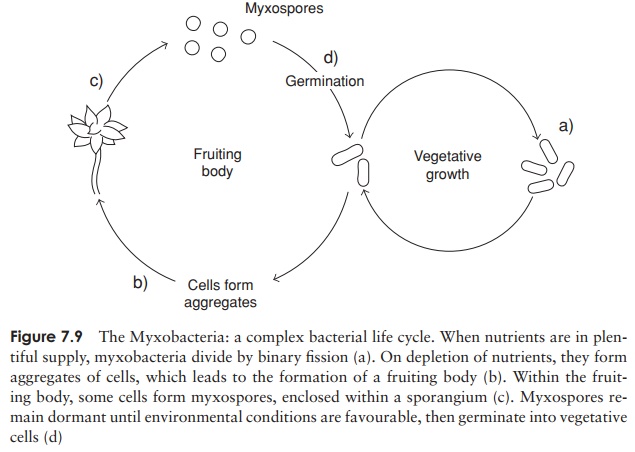
Another group of bacteria that may be regarded as predatory are the Myxobacteria (Figure 7.9). These are rod-shaped bacteria lacking flagella, which yet are motile by gliding along a solid surface, aided by the excretion of extracellular polysaccharides. For this reason they are sometimes referred to as the gliding bacteria. They are heterotrophs, typically requiring complex organic nutrients, which they obtain by the lysis of other types of bacteria. Thus, unlike Bdellovibrio, they digest their prey before they ingest it. When a rich supply of nutrients is not available, many thousands of cells may aggregate to form fruiting bodies, inside which myxospores develop. These are able to resist drought and lack of nutrients for many years. Myxobacteria exhibit the most complex life cycles of any procaryote so far studied.
Representative genera: Myxococcus, Chondromyces
Spirilla
Collected together under this heading are several genera of aerobic (mostly mi-croaerophilic) spiral-shaped bacteria with polar flagella. These include free-living, sym-biotic and parasitic types.
Spirilla such as Aquaspirillum and Magnetospirillum contain magnetosomes, intra-cellular particles of iron oxide (magnetite, Fe3O4). Such magnetotactic bacteria have the remarkable ability to orientate themselves with respect to the earthŌĆÖs magnetic field (magnetotaxis).
Two important pathogens of humans are included in the spirilla; Campylobacterjejuni is responsible for foodborne gastroenteritis, while Helicobacter pylori has in recenttimes been identified as the cause of many cases of peptic ulcers.
Representative genera: Magnetospirillum, Campylobacter
Rickettsia
This group comprises arthropod-borne intracellular parasites of vertebrates, and in-cludes the causative agents of human diseases such as typhus and Rocky Mountainspotted fever. The bacteria are taken up by host phagocytic cells, where they multiplyand eventually cause lysis.
The Rickettsia are aerobic organotrophs, but some possess an unusual mode of en-ergy metabolism, only being able to oxidise intermediate metabolites such as glutamate and succinate, which they obtain from their host. Rickettsia and Coxiella, the two main genera, are not closely related phylogenetically and are placed in the ╬▒- and ╬│ - Proteobacteria, respectively.
Representative genera: Rickettsia, Coxiella
Neisseria and related Proteobacteria
All members of this loose collection of bacteria are aerobic non-motile cocci, typically seen as pairs, with flattened sides where they join. Some however only assume this morphology during stationary growth phase. Many are found in warm-blooded animals, and some species are pathogenic. The genus Neisseria includes species responsible for gonorrhoea and meningitis in humans.
Other Gram-negative phyla
The following section considers those Gram-negative bacteria not included in the Pro-teobacteria. These phyla are not closely related in the phylogenetic sense, either to each other or to the Proteobacteria.
Related Topics