Chapter: Essential Microbiology: Procaryote Diversity
Domain: Archaea : Classification and General features
Domain: Archaea
Studies on 16S ribosomal RNA sequences by Carl Woese and colleagues allowed the construction of phylogenetic trees for the procaryotes, showing their evolutionary relatedness. Figure 7.1 shows how the major procaryotic groups are thought to be related, based on 16S rRNA data. The work of Woese also revealed that one group of procaryotes differed from all the others. As described, the Archaea are now regarded as being quitedistinct from the Bacteria (sometimes called Eubacteria). Together with the Eucarya, these form the three domains of life (Figure 3.1). As can be seen in Table 7.1, archaea
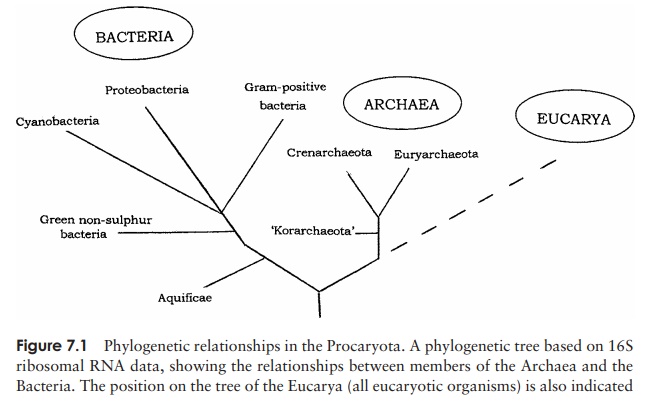
share some features in common with other bacteria and some with eucaryotes. Extending nucleic acid analysis to other genes has shown that members of the Archaea pos-sess many genes not found in any other type of bacteria.
General features of the Archaea
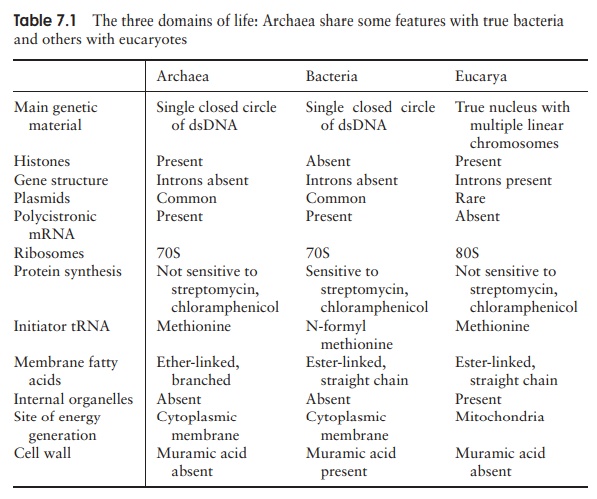
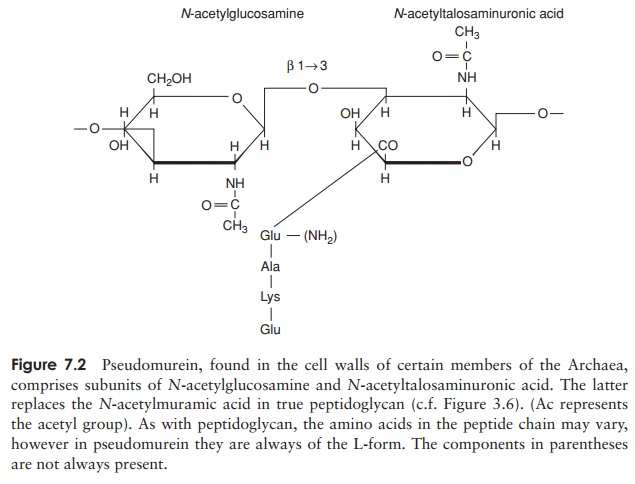
Members of the Archaea show considerable diversity of both morphology and physiol-ogy. In view of the fact that the Archaea remained unidentified as a separate group for so many years, it should come as no surprise that they do not display any obvious morpho-logical differences from true bacteria, and all the main cell shapes are rep-resented. More unusual shapes are also encountered in archaea; members of the genus Haloarcula have flattened square or triangular cells! Both Gram-positive and Gram-negative forms of archaea are found, but neither possesses true peptidoglycan. Some types have a so-called pseudomurein, composed of different substituted polysaccharides and L-amino acids (Figure 7.2). Most archaea, however, have cell walls composed of a layer of proteinaceous subunits known as an S-layer, directly associated with the cell membrane. This difference in cell wall chemistry means that members of the Archaea are not susceptible to antibacterial agents such as lysozyme and penicillin, whose action is directed specifically towards peptidoglycan. Differences are also found in the make-up
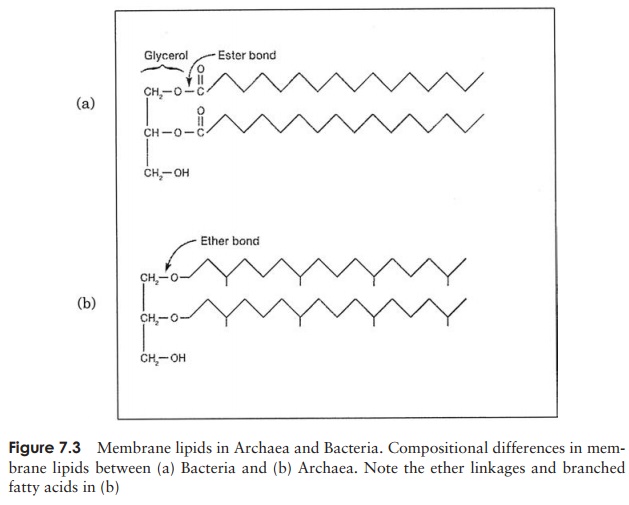
of archaean membranes, where the lipid component of membranes contains branched isoprenes instead of fatty acids, and these are joined to glycerol by ether-linkages, rather than the ester-linkages found in true bacteria (Figure 7.3). The diversity of archaea ex-tends into their adopted means of nutrition and metabolism: aerobic/anaerobic and autotrophic/heterotrophic forms are known. Many members of the Archaea are found in extreme environments such as deep-sea thermal vents and salt ponds. Some extreme thermophiles are able to grow at temperatures well over 100 ŌŚ” C, while psychrophilic forms constitute a substantial proportion of the microbial population of Antarctica. Similarly, examples are to be found of archaea that are active at extremes of acidity, alkalinity or salinity. Initially it was felt that archaea were limited to such environments because there they faced little competition from true bacteria or eucaryotes. Recent stud-ies have shown however that archaea are more widespread in their distribution, making up a significant proportion of the bacterial biomass found in the worldŌĆÖs oceans, and also being found in terrestrial and semiterrestrial niches. The reason that this lay unde-tected for so long is that these organisms cannot as yet be cultured in the laboratory, and their presence can only be inferred by the use of modern DNA-based analysis.
Classification of the Archaea
According to the second edition of Bergey, the Archaea are divided into two phyla, the Euryarchaeota and Crenarchaeota.
A third phylum, the Korarchaeota has been proposed, whose members are known only from molecular studies, while the recent discovery of Nanoarchaeum equitans has led to the proposal of yet another archaean phylum. Countless more species of archaea are thought to exist, which like the Korarchaeota, have not yet been successfully cultured in the laboratory.
The phylum Eurychaeota is a bigger group than the Crenarchaeota, and includes halophilic and methanogenic forms. The former are aerobic heterotrophs, requiring a chloride concentration of at least 1.5 M (generally 2.0ŌĆō4.0 M) for growth. One species, Halobacterium salinarum, is able to carry out a unique form of photosynthesis usingthe bacterial pigment bacteriorhodopsin, and uses the ATP so generated for the active transport into the cell of the chloride ions it requires.
Members of the Euryarchaeota such as Methanococcus and Methanobacterium are unique among all life forms in their ability to generate methane from simple carbon com-pounds. They are strict anaerobes found in environments such as hot springs, marshes and the gut of ruminant mammals. The methane is derived from the metabolism of var-ious simple carbon compounds such as carbon dioxide or methanol in reactions linked to the production of ATP. e.g.
CO2+4H2ŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌåÆ CH4+ 2H2O
CH3OH +H2ŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌåÆ CH4+ H2O
In addition, a few species can cleave acetate to produce methane:
CH3COOŌłÆ+ H2O ŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌłÆŌåÆ CH4+ HCO3ŌłÆ
his acetotrophic reaction is responsible for the much of the methane production in sewage sludges. Although sharing the unique facility to generate methane, some of the morphic. A unique membrane lipid composition allows them to withstand temperatures of well over 50 ŌŚ” C. Thermococci are anaerobic extreme thermophiles found in anoxic thermal waters at temperatures as high as 95 ŌŚ” C. Enzymes isolated from thermococci have found a variety of applications. A thermostable DNA polymerase fromPyrococcusfuriosus is used as an alternative to Taq polymerase in the polymerase chain reaction (PCR).
Members of the Crenarchaeota are nearly all extreme thermophiles, many of them capable of growth at temperatures in excess of 100 ŌŚ” C, including Pyrolobus fumarii,which has an optimum growth temperature of 106 ŌŚ” C, and can survive autoclaving at 121 ŌŚ” C.
Many utilise inorganic sulphur compounds as either a source or acceptor of electrons (respectively, oxidation to H2SO4 or reduction to H2S).
Crenarchaeotes are mostly anaerobic, and are thought by many to resemble the common ancestors of all bacteria.
Representative genera: Thermoproteus, Sulfolobus
Related Topics