Chapter: Modern Pharmacology with Clinical Applications: Ganglionic Blocking Drugs and Nicotine
Ganglionic Transmission
GANGLIONIC
TRANSMISSION
Transmission through
autonomic ganglia is more com-plex than neurotransmission at the neuromuscular
and postganglionic neuroeffector junctions and is subject to numerous
pharmacological and physiological influ-ences. In some ganglionic synapses,
especially at parasympathetic ganglia, there is a simple presynaptic to postsynaptic
cell relationship; in others, the presyn-aptic to postsynaptic cell
relationship may involve neu-rons interposed between the presynaptic and
postsy-naptic elements (interneurons).
In a variety of sympathetic
and certain parasympa-thetic ganglion cells (e.g., vagal ganglia in the
sinoatrial node), cells exhibiting the characteristic catecholamine
fluorescence spectrum have been found. These cells are referred to as small
intensely fluorescent (SIF) cells. At some autonomic ganglia, the SIF cell is a
true interneu-ron, receiving afferent innervation from preganglionic
cholinergic neurons and forming efferent synapses with postganglionic neurons.
At other autonomic ganglia, its function is not completely understood, but the
SIF cell is believed to play a role in the modulation of ganglionic
transmission. Many SIF cells are thought to contain dopamine or norepinephrine
as their neurotransmitter.
Unlike the receptors at
postganglionic neuroeffector junctions or at skeletal neuromuscular junctions,
both types of cholinergic receptors, that is, nicotinic and mus-carinic, are
present on the cell bodies of the postgan-glionic neurons. Stimulation of the
preganglionic neuron results in the release of acetylcholine (ACh) from the
preganglionic nerve terminal, which in turn activates postganglionic
cholinergic receptors and leads ulti-mately to the formation of a propagated
action potential down the postganglionic axon. At the more complicated
synapses, the release of ACh from preganglionic neurons results in the appearance
of complex postsynaptic po-tential changes consisting of several temporally
ar-ranged components. There is an initial fast excitatory postsynaptic
potential (EPSP) followed by a succession of much slower postsynaptic potential
changes, including a slow EPSP that lasts for 2 to 5 seconds, a slow
in-hibitory postsynaptic potential (IPSP) lasting about 10 seconds, and a late
slow EPSP lasting for 1 to 2 minutes.
There is considerable
diversity among nicotinic acetylcholine receptors, and at least one source of
this diversity is the multiplicity of acetylcholine receptor genes.
Cholinergic–nicotinic receptors in skeletal mus-cle are different from those in
autonomic ganglia and the central nervous system.
Excitatory and Inhibitory Potentials
The interaction of ACh with
the postsynaptic nicotinic receptor results in depolarization of the membrane,
an influx of NA+ and Ca++ through a neuronal nicotinic
re-ceptor channel, and the generation of the fast EPSP. This change in
postsynaptic potential is principally re-sponsible for the generation of the
propagated action potential in the postganglionic neuron. Generally, sev-eral
presynaptic terminals innervate a single ganglion cell, and several
preganglionic axon terminals must fire simultaneously for transmission to take
place. Ganglionic blocking agents prevent transmission by in-terfering with the
postsynaptic action of ACh. The drugs either interact with the
nicotinic–cholinergic receptor itself or with the associated ionic channel
complex.
Interaction of ACh with the
postsynaptic ganglionic cell muscarinic receptor is responsible for slowly
devel-oping depolarization, the slow EPSP, which has a longer latency than the
fast EPSP and a duration of 30 to 60 seconds. The slow EPSP is due to inhibition
of a voltage-dependent K+ current called the M current, and
inhibi-tion of the M current involves activation of G proteins. At least five
types of muscarinic receptors (M1, M2, M3, M4
and M5) have been identified using functional stud-ies and at least
five subtypes (m1, m2, m3, m4, and
m5) identified by molecular cloning techniques. The M1
re-ceptor, which appears responsible for inhibiting the M current, can be
blocked by atropine.
Release of ACh may activate
SIF cells between pre-ganglionic and postganglionic neurons. In this case,
acti-vation of a muscarinic receptor on the SIF cells results in the release of
a catecholamine; this in turn activates a receptor on the postganglionic cell,
leading to the slow IPSP. The catecholamine most frequently released from SIF
cells appears to be dopamine. Finally, a late slow EPSP, lasting for 1 to 2
minutes, can be seen at some ganglionic synapses. The mediator is unclear, but
it is now well established that there are a large number of peptides in the
ganglia, including luteinizing hormone– releasing hormone (LHRH), substance P,
angiotensin, calcitonin gene related peptide, vasoactive intestinal
polypeptide, neuropeptide Y, and enkephalin.
In addition to the
cholinergic and adrenergic recep-tors on autonomic ganglion cells, there also
appear to be receptors for a variety of excitatory and inhibitory sub-stances,
including angiotensin, bradykinin, histamine, 5-hydroxytryptaimine (serotonin),
and substance P. Theexistence of these receptors provides a wide variety of options
to modulate ganglionic transmission. Agonists for these receptors most likely
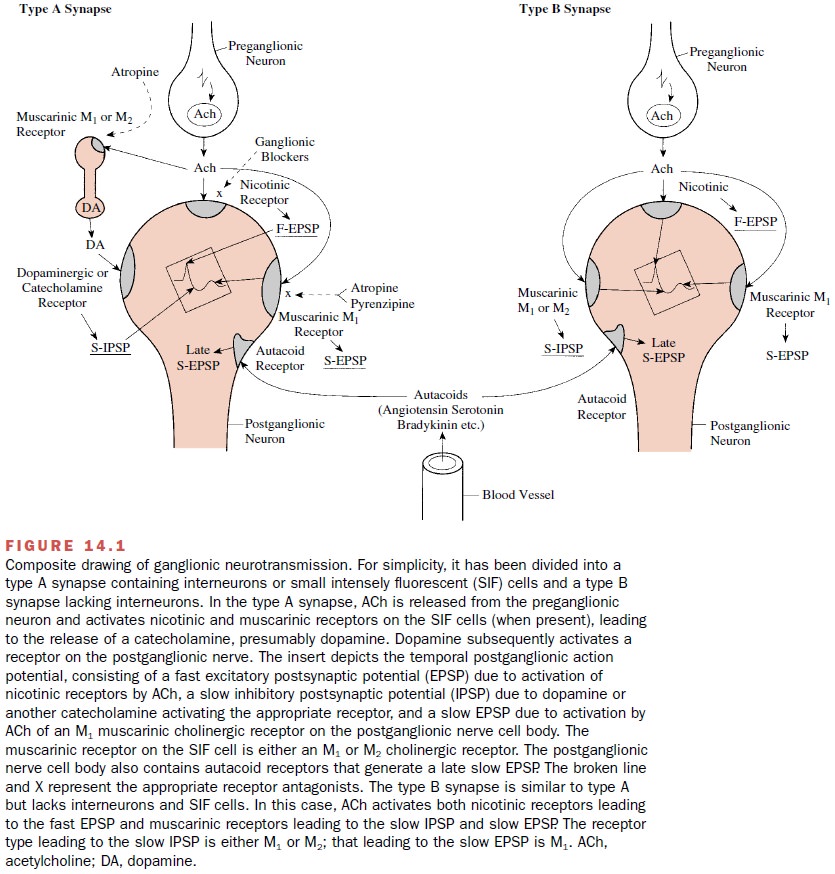
reach the ganglia through the circulation. A composite picture of the status of
gan-glionic transmission is shown in Figure 14.1. For simplic-ity, the figure
has been divided into a type A synapse, which includes SIF cells, and a type B
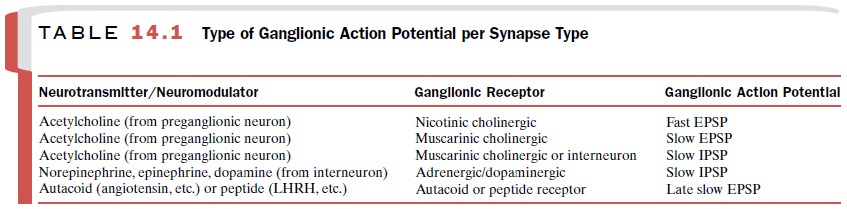
synapse, which lacks SIF cells. Table 14.1 summarizes the type of gan-glionic
action potential generated at various synapses, the type of receptor mediating
the response, and the pri-mary transmitter or mediator that activates the
receptor.
Related Topics