Chapter: Medical Physiology: The Sense of Hearing
Function of the Organ of Corti
Function of the Organ of Corti
The organ of Corti, shown in Figures 52–2, 52–3, and 52–7, is the receptor organ that generates nerve impulses in response to vibration of the basilar mem-brane. Note that the organ of Corti lies on the surface of the basilar fibers and basilar membrane. The actual sensory receptors in the organ of Corti are two spe-cialized types of nerve cells called hair cells—a single row of internal (or “inner”) hair cells, numbering about 3500 and measuring about 12 micrometers in diame-ter, and three or four rows of external (or “outer”) haircells, numbering about 12,000 and having diametersof only about 8 micrometers. The bases and sides of the hair cells synapse with a network of cochlea nerve endings. Between 90 and 95 per cent of these endings terminate on the inner hair cells, which emphasizes their special importance for the detection of sound.
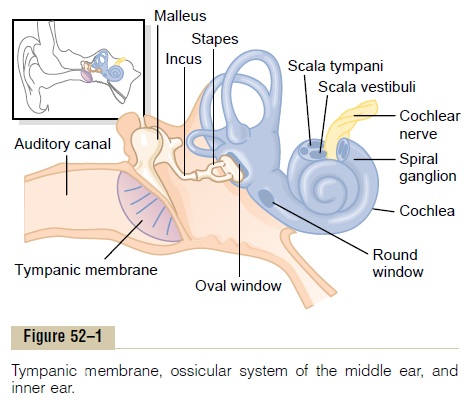
The nerve fibers stimulated by the hair cells lead to the spiral ganglion of Corti, which lies in the modiolus (center) of the cochlea. The spiral ganglion neuronal cells send axons—a total of about 30,000—into the cochlear nerve and then into the central nervoussystem at the level of the upper medulla. The relation of the organ of Corti to the spiral ganglion and to the cochlear nerve is shown in Figure 52–2.
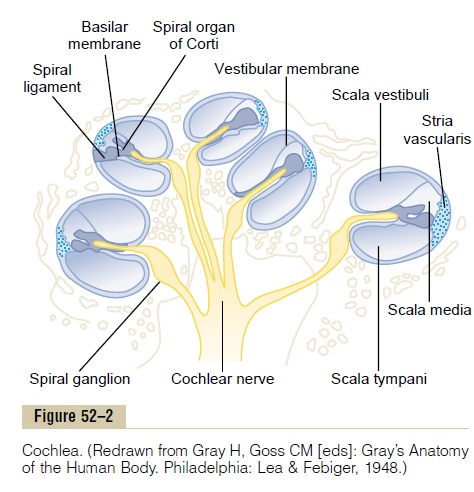
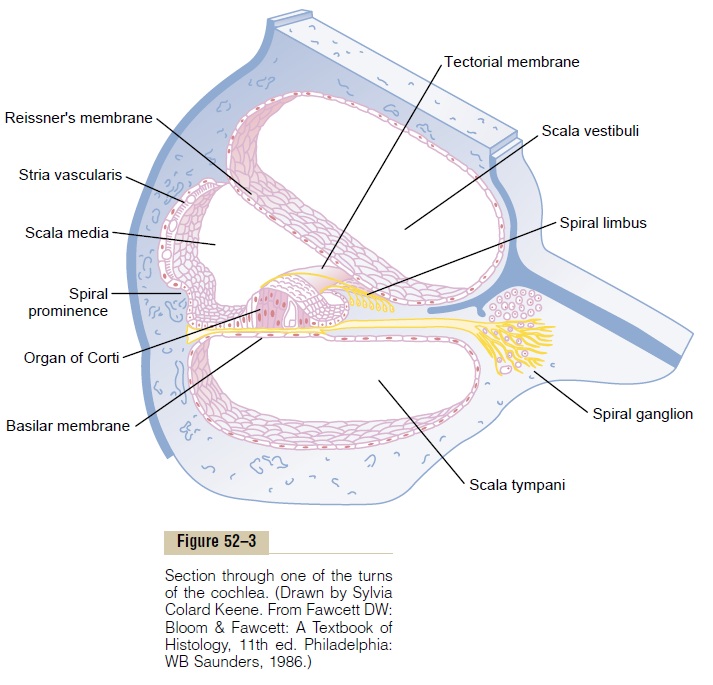
Excitation of the Hair Cells. Note in Figure 52–7 thatminute hairs, or stereocilia, project upward from the hair cells and either touch or are embedded in the surface gel coating of the tectorial membrane, which lies above the stereocilia in the scala media. These hair cells are similar to the hair cells found in the macula and cristae ampullaris of the vestibular apparatus. Bending of the hairs in one direction depolarizes the hair cells, and bending in the opposite direction hyperpolarizes them. This in turn excites the auditory nerve fibers synaps-ing with their bases.
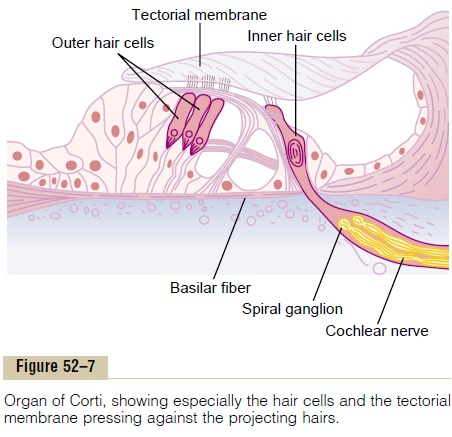
Figure 52–8 shows the mechanism by which vibra-tion of the basilar membrane excites the hair endings. The outer ends of the hair cells are fixed tightly in a rigid structure composed of a flat plate, called the retic-ular lamina, supported by triangular rods of Corti,which are attached tightly to the basilar fibers. The basilar fibers, the rods of Corti, and the reticular lamina move as a rigid unit.
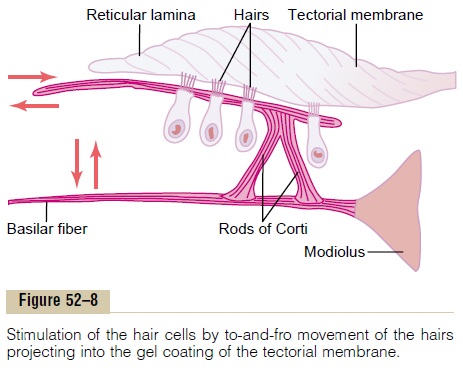
Upward movement of the basilar fiber rocks the reticular lamina upward and inward toward the modi-olus. Then, when the basilar membrane moves down-ward, the reticular lamina rocks downward and outward. The inward and outward motion causes thehairs on the hair cells to shear back and forth against the tectorial membrane. Thus, the hair cells are excited whenever the basilar membrane vibrates.
Auditory Signals Are Transmitted Mainly by the Inner Hair Cells.
Even though there are three to four times as many outer hair cells as inner hair cells, about 90 per cent of the auditory nerve fibers are stimulated by the inner cells rather than by the outer cells. Yet, despite this, if the outer cells are damaged while the inner cells remain fully functional, a large amount of hearing loss occurs. Therefore, it has been proposed that the outer hair cells in some way control the sensitivity of the inner hair cells at different sound pitches, a phenomenon called “tuning” of the receptor system.
In support of this concept, a large number of retro-grade nerve fibers pass from the brain stem to the vicinity of the outer hair cells. Stimulating these nerve fibers can actually cause shortening of the outer hair cells and possibly also change their degree of stiffness. These effects suggest a retrograde nervous mechanism for control of the ear’s sensitivity to different sound pitches, activated through the outer hair cells.
Hair Cell Receptor Potentials and Excitation of Auditory Nerve Fibers. The stereocilia (the “hairs” protruding from theends of the hair cells) are stiff structures because each has a rigid protein framework. Each hair cell has about 100 stereocilia on its apical border. These become pro-gressively longer on the side of the hair cell away from the modiolus, and the tops of the shorter stereocilia are attached by thin filaments to the back sides of their adjacent longer stereocilia. Therefore, whenever the cilia are bent in the direction of the longer ones, the tips of the smaller stereocilia are tugged outward from the surface of the hair cell. This causes a mechanical transduction that opens 200 to 300 cation-conducting channels, allowing rapid movement of positively charged potassium ions from the surrounding scala media fluid into the stereocilia, which causes depolar-ization of the hair cell membrane.
Thus, when the basilar fibers bend toward the scala vestibuli, the hair cells depolarize, and in the opposite direction they hyperpolarize, thereby generating an alternating hair cell receptor potential. This, in turn, stimulates the cochlear nerve endings that synapse with the bases of the hair cells. It is believed that a rapidly acting neurotransmitter is released by the hair cells at these synapses during depolarization. It is pos-sible that the transmitter substance is glutamate, but this is not certain.
Endocochlear Potential. To explain even more fully theelectrical potentials generated by the hair cells, we need to explain another electrical phenomenon called the endocochlear potential: The scala media is filled with afluid called endolymph, in contradistinction to the peri-lymph present in the scala vestibuli and scala tympani.The scala vestibuli and scala tympani communicate directly with the subarachnoid space around the brain, so that the perilymph is almost identical with cere-brospinal fluid. Conversely, the endolymph that fills the scala media is an entirely different fluid secreted by the stria vascularis, a highly vascular area on the outer wall of the scala media. Endolymph contains a high concentration of potassium and a low concentration of sodium, which is exactly opposite to the contents of perilymph.
An electrical potential of about +80 millivolts exists all the time between endolymph and perilymph, with positivity inside the scala media and negativity outside. This is called the endocochlear potential, and it is gen-erated by continual secretion of positive potassium ions into the scala media by the stria vascularis.
The importance of the endocochlear potential is that the tops of the hair cells project through the reticular lamina and are bathed by the endolymph of the scala media, whereas perilymph bathes the lower bodies of the hair cells. Furthermore, the hair cells have a nega-tive intracellular potential of –70 millivolts with respect to the perilymph but –150 millivolts with respect to the endolymph at their upper surfaces where the hairs project through the reticular lamina and into the endolymph. It is believed that this high electrical poten-tial at the tips of the stereocilia sensitizes the cell an extra amount, thereby increasing its ability to respond to the slightest sound.
Related Topics