Chapter: Medical Physiology: The Sense of Hearing
Function of the Cerebral Cortex in Hearing
Function of the Cerebral Cortex in Hearing
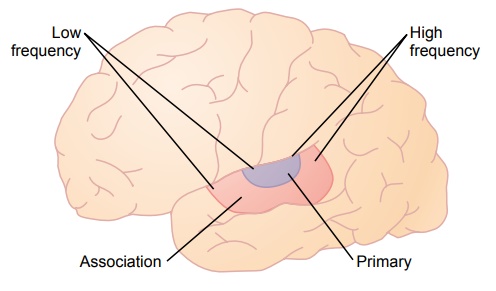
The projection area of auditory signals to the cerebral cortex is shown in Figure 52–11, which demonstrates that the auditory cortex lies principally on the supratemporal plane of the superior temporal gyrus butalso extends onto the lateral side of the temporal lobe,over much of the insular cortex, and even onto the lateral portion of the parietal operculum.
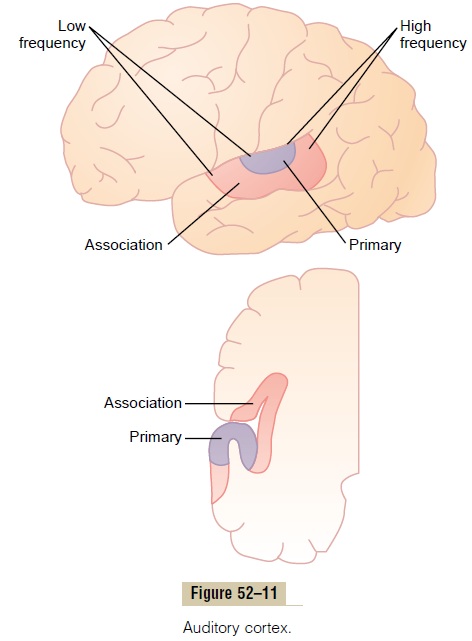
Two separate subdivisions are shown in Figure 52–11: the primary auditory cortex and the auditoryassociation cortex (also called the secondary auditory cortex). The primary auditory cortex is directly excitedby projections from the medial geniculate body, whereas the auditory association areas are excited secondarily by impulses from the primary auditory cortex as well as by some projections from thalamic association areas adjacent to the medial geniculate body.
Sound Frequency Perception in the Primary Auditory Cortex.
At least six tonotopic maps have been found in the primary auditory cortex and auditory association areas. In each of these maps, high-frequency sounds excite neurons at one end of the map, whereas low-frequency sounds excite neurons at the opposite end. In most, the low-frequency sounds are located anteri-orly, as shown in Figure 52–11, and the high-frequency sounds are located posteriorly. This is not true for all the maps.
The question one must ask is, Why does the auditory cortex have so many different tonotopic maps? The answer, presumably, is that each of the sep-arate areas dissects out some specific feature of the sounds. For instance, one of the large maps in the primary auditory cortex almost certainly discriminates the sound frequencies themselves and gives the person the psychic sensation of sound pitches. Another map is probably used to detect the direction from which the sound comes. Other auditory cortex areas detect special qualities, such as the sudden onset of sounds, or perhaps special modulations, such as noise versus pure frequency sounds.
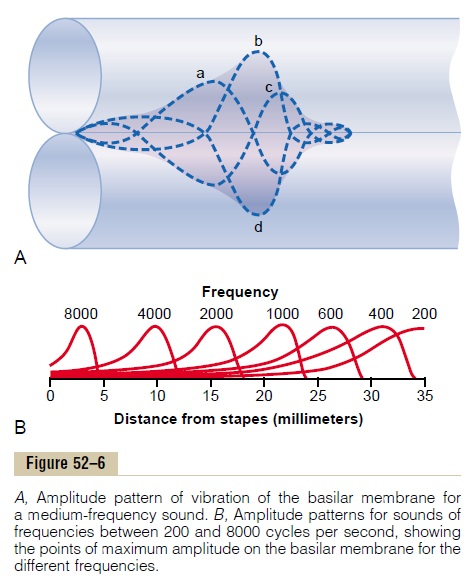
The frequency range to which each individual neuron in the auditory cortex responds is much narrower than that in the cochlear and brain stem relay nuclei. Referring back to Figure 52–6B, note that the basilar membrane near the base of the cochlea is stimulated by sounds of all frequencies, and in the cochlear nuclei, this same breadth of sound represen-tation is found. Yet, by the time the excitation has reached the cerebral cortex, most sound-responsive neurons respond to only a narrow range of frequen-cies rather than to a broad range. Therefore, some-where along the pathway, processing mechanisms “sharpen” the frequency response. It is believed that this sharpening effect is caused mainly by the phe-nomenon of lateral inhibition, which is discussed in relation to mechanisms for transmit-ting information in nerves. That is, stimulation of the cochlea at one frequency inhibits sound frequencies on both sides of this primary frequency; this is caused by collateral fibers angling off the primary signal pathway and exerting inhibitory influences on adjacent pathways. The same effect has been demon-strated to be important in sharpening patterns of somesthetic images, visual images, and other types of sensations.
Many of the neurons in the auditory cortex, espe-cially in the auditory association cortex, do not respondonly to specific sound frequencies in the ear. It is believed that these neurons “associate” different sound frequencies with one another or associate sound information with information from other sensory areas of the cortex. Indeed, the parietal portion of the auditory association cortex partly overlaps somatosen-sory area II, which could provide an easy opportunity for the association of auditory information with somatosensory information.
Discrimination of Sound “Patterns” by the Auditory Cortex.
Complete bilateral removal of the auditory cortex does not prevent a cat or monkey from detecting sounds or reacting in a crude manner to sounds. However, it does greatly reduce or sometimes even abolish the animal’s ability to discriminate dif-ferent sound pitches and especially patterns of sound. For instance, an animal that has been trained to rec-ognize a combination or sequence of tones, one fol-lowing the other in a particular pattern, loses this ability when the auditory cortex is destroyed; further-more, the animal cannot relearn this type of response. Therefore, the auditory cortex is especially important in the discrimination of tonal andsequential soundpatterns.
Destruction of both primary auditory cortices in the human being greatly reduces one’s sensitivity for hearing. Destruction of one side only slightly reduces hearing in the opposite ear; it does not cause deafness in the ear because of many crossover con-nections from side to side in the auditory neural pathway. However, it does affect one’s ability to localize the source of a sound, because comparative signals in both cortices are required for the localiza-tion function.
Lesions that affect the auditory association areas but not the primary auditory cortex do not decrease a person’s ability to hear and differentiate sound tones or even to interpret at least simple patterns of sound. However, he or she is often unable to interpret themeaning of the sound heard. For instance, lesions inthe posterior portion of the superior temporal gyrus, which is called Wernicke’s area and is part of the audi-tory association cortex, often make it impossible for a person to interpret the meanings of words even though he or she hears them perfectly well and can even repeat them.
Related Topics