Chapter: Medical Physiology: Organization of the Nervous System, Basic Functions of Synapses, “Transmitter Substances”
Electrical Events During Neuronal Inhibition - Central Nervous System Synapses
Electrical Events During Neuronal Inhibition
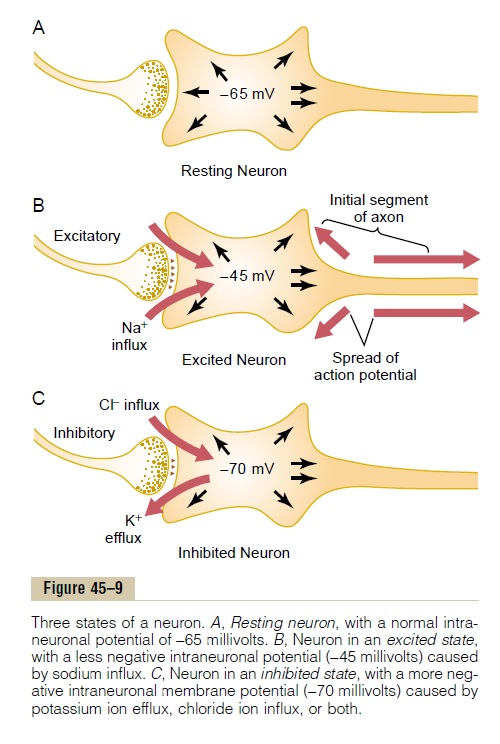
Effect of Inhibitory Synapses on the Postsynaptic Membrane— Inhibitory Postsynaptic Potential. The inhibitory synapsesopen mainly chloride channels, allowing easy passageof chloride ions. Now, to understand how the inhibitory synapses inhibit the postsynaptic neuron, we must recall what we learned about the Nernst potential for chloride ions. We calculated the Nernst potential for chloride ions to be about -70 millivolts. This potential is more negative than the -65 millivolts normally present inside the resting neuronal mem-brane. Therefore, opening the chloride channels will allow negatively charged chloride ions to move from the extracellular fluid to the interior, which will make the interior membrane potential more negative than normal, approaching the -70 millivolt level.
Opening potassium channels will allow positively charged potassium ions to move to the exterior, and this will also make the interior membrane potential more negative than usual. Thus, both chloride influx and potassium efflux increase the degree of intracel-lular negativity, which is called hyperpolarization. This inhibits the neuron because the membrane potential is even more negative than the normal intracellular potential. Therefore, an increase in negativity beyond the normal resting membrane potential level is called aninhibitory postsynaptic potential (IPSP).
Figure 45–9C shows the effect on the membrane potential caused by activation of inhibitory synapses, allowing chloride influx into the cell and/or potassium efflux out of the cell, with the membrane potential decreasing from its normal value of -65 millivolts to the more negative value of -70 millivolts. This mem-brane potential is 5 millivolts more negative than normal and is therefore an IPSP of -5 millivolts, which inhibits transmission of the nerve signal through the synapse.
Presynaptic Inhibition
In addition to inhibition caused by inhibitory synapses operating at the neuronal membrane, which is called postsynaptic inhibition, another type of inhibitionoften occurs at the presynaptic terminals before the signal ever reaches the synapse. This type of inhibition, called presynaptic inhibition, occurs in the following way.
Presynaptic inhibition is caused by release of an inhibitory substance onto the outsides of the presy-naptic nerve fibrils before their own endings terminate on the postsynaptic neuron. In most instances, the inhibitory transmitter substance is GABA (gamma-aminobutyric acid). This has a specific effect of opening anion channels, allowing large numbers of chloride ions to diffuse into the terminal fibril. The negative charges of these ions inhibit synaptic trans-mission because they cancel much of the excitatory effect of the positively charged sodium ions that also enter the terminal fibrils when an action potential arrives.
Presynaptic inhibition occurs in many of the sensory pathways in the nervous system. In fact, adjacent sensory nerve fibers often mutually inhibit one another, which minimizes sideways spread and mixing of signals in sensory tracts. We discuss the importance of this phenomenon more fully in subsequent.
Time Course of Postsynaptic Potentials
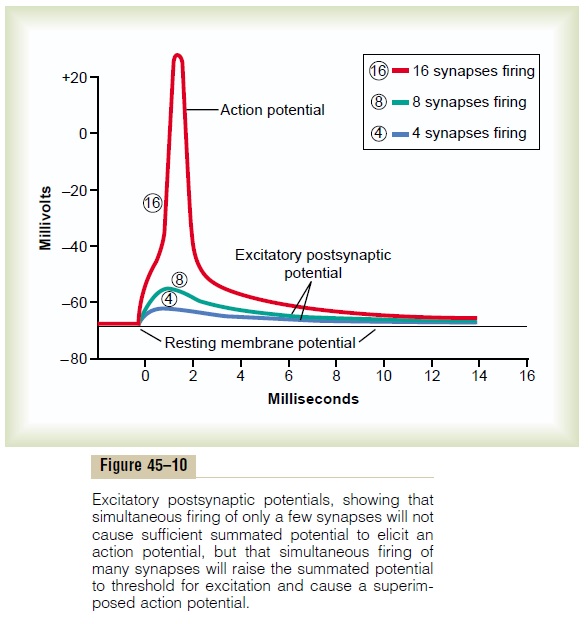
When an excitatory synapse excites the anterior motor neuron, the neuronal membrane becomes highly per-meable to sodium ions for 1 to 2 milliseconds. During this very short time, enough sodium ions diffuse rapidly to the interior of the postsynaptic motor neuron to increase its intraneuronal potential by a few millivolts, thus creating the excitatory postsynaptic potential (EPSP) shown by the blue and green curves of Figure 45–10. This potential then slowly declines over the next 15 milliseconds because this is the time required for the excess positive charges to leak out of the excited neuron and to re-establish the normal resting membrane potential.
Precisely the opposite effect occurs for an IPSP; that is, the inhibitory synapse increases the permeability of the membrane to potassium or chloride ions, or both, for 1 to 2 milliseconds, and this decreases the intra-neuronal potential to a more negative value than normal, thereby creating the IPSP. This potential also dies away in about 15 milliseconds.
Other types of transmitter substances can excite or inhibit the postsynaptic neuron for much longer periods—for hundreds of milliseconds or even for seconds, minutes, or hours. This is especially true for some of the neuropeptide types of transmitter substances.
“Spatial Summation” in Neurons— Threshold for Firing Excitation of a single presynaptic terminal on the surface of a neuron almost never excites the neuron. The reason for this is that sufficient transmitter sub-stance is released by a single terminal to cause an EPSP usually no greater than 0.5 to 1 millivolt, instead of the 10 to 20 millivolts normally required to reach threshold for excitation. However, many presynaptic terminals are usually stimulated at the same time. Even though these terminals are spread over wide areas of the neuron, their effects can still summate; that is, they can add to one another until neuronal exci-tation does occur. The reason for this is the following: It was pointed out earlier that a change in potential at any single point within the soma will cause the poten-tial to change everywhere inside the soma almost exactly equally. This is true because of the very high electrical conductivity inside the large neuronal cell body. Therefore, for each excitatory synapse that dis-charges simultaneously, the total intrasomal potential becomes more positive by 0.5 to 1.0 millivolt.
When the EPSP becomes great enough, the threshold forfiring will be reached and an action potential willdevelop spontaneously in the initial segment of the axon. This is demonstrated in Figure 45–10. The bottom postsynaptic potential in the figure was caused by simultaneous stimulation of 4 synapses; the next higher potential was caused by stimulation of 8 synapses; finally, a still higher EPSP was caused by stimulation of 16 synapses. In this last instance, the firing threshold had been reached, and an action potential was generated in the axon.
This effect of summing simultaneous postsynaptic potentials by activating multiple terminals on widely spaced areas of the neuronal membrane is called spatial summation.
“Temporal Summation”
Each time a presynaptic terminal fires, the released transmitter substance opens the membrane channels for at most a millisecond or so. But the changed post-synaptic potential lasts up to 15 milliseconds after the synaptic membrane channels have already closed. Therefore, a second opening of the same channels can increase the postsynaptic potential to a still greater level, and the more rapid the rate of stimulation, the greater the postsynaptic potential becomes. Thus, suc-cessive discharges from a single presynaptic terminal, if they occur rapidly enough, can add to one another; that is, they can “summate.” This type of summation is called temporal summation.
Simultaneous Summation of Inhibitory and Excitatory Postsy-naptic Potentials. If an IPSP is tending todecreasethemembrane potential to a more negative value while an EPSP is tending to increase the potential at the same time, these two effects can either completely or par-tially nullify each other. Thus, if a neuron is being excited by an EPSP, an inhibitory signal from another source can often reduce the postsynaptic potential to less than threshold value for excitation, thus turning off the activity of the neuron.
“Facilitation” of Neurons
Often the summated postsynaptic potential is excita-tory but has not risen high enough to reach the thresh-old for firing by the postsynaptic neuron. When this happens, the neuron is said to be facilitated. That is, its membrane potential is nearer the threshold for firing than normal, but not yet at the firing level. Conse-quently, another excitatory signal entering the neuron from some other source can then excite the neuron very easily. Diffuse signals in the nervous system often do facilitate large groups of neurons so that they can respond quickly and easily to signals arriving from other sources.
Related Topics