Chapter: Health Management in Aquaculture: Immunity and biological methods of disease prevention and control
Crustacean immune system
THE CRUSTACEAN IMMUNE SYSTEM
The crustacean immune system lacks the immune memory characteristic of vertebrates and have to rely on the innate or non-specific response. However, crustaceans are no different from the vertebrates in that their immune defense is largely brought about by the activities of specialized blood cells or hemocytes. These crustacean hemocytes carry out phagocytosis, encapsula-tion, produce antimicrobial substances to remove or neutralize foreign par-ticles and infectious agents.
Hemocyte types
Based on their morphological appearance and their staining properties, differ-ent hemocyte types can be distinguished in crustacean blood. However, these hemocyte types do not fall into the same groups as those found in fish and other vertebrates. The three types of hemocytes that can be isolated from crus-tacean hemolymph are the hyaline, the semigranular, and the granular (see Table 9 –2 for summary of their functions).
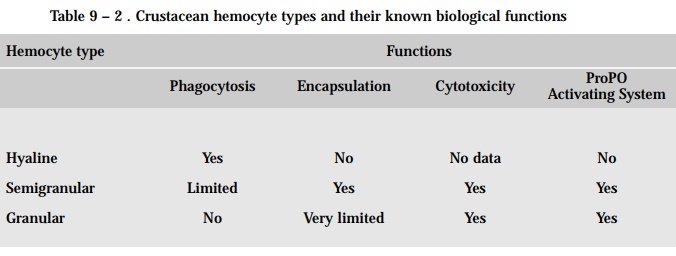
The hyaline hemocytes have phagocytic ability. This cell type, which lacks granules, is found in decapod crustaceans, but apparently the relative number of this cell type vary considerably among species.
The semigranular hemocytes are characterized by the presence of a number of small granules in their cytoplasm making them resemble the granulocytes of vertebrates. These cells respond to microbial cell wall polysaccharides such as bacterial lipopolysaccharides (LPS) and the b-1,3 glucans of fungi. They also have the ability to encapsulate foreign particles.
The granular hemocytes are characterized by the presence of large vesicles or granules in their cytoplasm. This suggests a role in the production, storage and secretion of antimicrobial compounds. They do not have phagocytic activity and their ability to encapsulate foreign particles is limited. The primary role of granular hemocytes is to store prophenol oxidase, which is key compound in crustacean defense reactions. These cells can be induced to unload and activate the prophenol oxidase by the presence of B-1,3 glucans, peptidoglycans and LPS from microorganisms. Once activated, the phenol oxidase catalyzes the oxidation of phenols to semiquinones and quinones that are highly toxic to microbes due to their high reactivity.
Defense Mechanisms in Crustaceans
Phagocytosis
Phagocytic cells are found throughout the animal kingdom. In lower invertebrates they also serve a nutritive function and in higher phyla they become more specialized by assuming a defensive role against microbial infections. A microorganism that has penetrated the exoskeleton and enters the tissues or the blood is immediately attacked by phagocytic cells that are specialized to engulf and digest particulate matter. In crustaceans, these are the hyaline hemocytes. Their primary function is to clear the body of foreign particles in-cluding virus, bacteria and fungal cells. Objects too large to be phagocytosed by one cell are trapped inside aggregates of hemocytes.
Nodule Formation and Encapsulation
When crustaceans are invaded by a large number of microorganisms that ex-ceed the capacity of phagocytic cells, nodule formation or cell clumping oc-curs. The microorganisms become entrapped in several layers of hemocytes, and generally the nodule become heavily melanized because of the host’s phenoloxidase activity.
When a parasite is too large to become engulfed by phagocytosis several hemocytes will then collaborate by sealing off the foreign particle from circula-tion. This process is known as encapsulation. The semigranular hemocytes are the first cell to react to foreign particles and to encapsulate any invading in-truder. However, little is known about the mechanisms by which foreign par-ticles or microbes are dealt with after being engulfed or encapsulated by hemocytes.
Cytotoxicity
Warm-blooded animals (higher vertebrates) produce a specialized white blood cell capable of killing tumor cells and cells infected by virus. Such cells are called natural killer cells. Among the white blood cells in the hemolymph of crustaceans there are populations of specialized hemocytes that, like the natu-ral killer cells in mammals, have the ability to kill foreign cells, tumor cells and non-tumor target cells.
Lectins
Agglutinating substances or lectins are present in the blood of a number of different crustaceans. Lectins are proteins or glycoproteins that have the ability to recognize and bind to the carbohydrates on the bacterial or fungal surfaces. Lectins do not have catalytic or enzymatic activity, their action is simply to immobilize or agglutinate microorganisms and then mediate the binding be-tween hemocyte surfaces and the microorganisms (or other foreign bodies) and thus function as an opsonin.
Antimicrobial Proteins or Peptides
The immune systems of arthropods also rely on the production of proteins and peptides that possess antimicrobial activity against a wide range of microor-ganisms. However, in crustaceans the presence and characterization of these antimicrobial peptides has been poorly studied until now. Recently, hemocytic proteins have been isolated in the crab, Carcinus maenas and a 6.5kDa antimi-crobial peptide has been characterized. In the penaeids, three antimicrobial peptides have been isolated from the hemocytes and plasma of Penaeusvanamei. They have been fully characterized and their cDNA cloned. Based on their biochemical and structural features, the three peptides do not belong to any group of peptides that have been hitherto described. The peptides were named penaeidins after the genus Penaeus. Studies on the role of the penaeidins in the immune system are still continuing, as are studies on the search and characterization of other antimicrobial peptides in shrimp.
The Clotting Reaction
Since crustaceans have an open circulatory system, wounds must be sealed immediately to stop blood loss and prevent the entry and distribution of mi-crobes within the body. In the shrimp the clotting process requires the presence of plasma proteins and cellular components. The key plasma protein that con-stitutes the clot has been named clotting protein or CP. It appears to be present in relatively high concentration in the hemolymph. The clotting reaction in crustaceans differs from that of vertebrates because aside from tissue damage as a trigger for the clotting cascade, the presence of microbial LPS is also a stimulus for the release of transglutaminase from hyaline cells that triggers the clotting process.
The Phenol Oxidase System
Parasites and microbes can gain entry into a crustacean body through wounds or as contaminants in the food. Some pathogens like the fungi penetrate the exoskeleton by secreting proteases and exerting mechanical forces. The re-sponse to this invasion can often be seen as dark spots in the cuticle and the intruders will become brown-black. The cause of this is melanin which is one of the end products of the phenoloxidase system. The enzyme responsible for the formation of melanin is phenoloxidase (PO). This enzyme (PO) catalyzes the oxidation of phenols to quinines that subsequently polymerize into mela-nin. During the formation of melanin, transient oxidation products are also formed which are highly reactive and toxic to microorganisms. The phenol oxi-dase system is thus an important component of the crustacean immune sys-tem.
A crucially significant feature of the phenol oxidase system is that it is able to identify a real infection and it can only be switched on by signals that are uniquely associated with the physical presence of pathogens. Crustaceans use the lipopolysaccharides LPS and the b-1,3 glucan molecules, which are compo-nents of the cell walls of microbes, as the specific signals to activate the phenol oxidase system. Crustacean hemolymph contains specialized binding proteins that seeks out and binds to the LPS and glucans of microbes. Once these bind-ing proteins have reacted with their target LPS and glucans, they then bind to a specific receptor on the hemocytes (both semigranular and granular) and in-duce degranulation and the release of the prophenol oxidase, which can be converted from its proform into the active enzyme (phenol oxidase) again upon contact with microbial LPS and glucans. These specific binding proteins, whose structures have already been elucidated, can also act as opsonins that stimulate phagocytosis. Thus the crustacean phenol oxidase system is exquis-itely designed to be specific against microbes and to avoid metabolically costly and harmful false alarms.
Related Topics