Chapter: Plant Anatomy:An Applied Approach: The stem
Stems - cross sectional appearance
Stems - cross sectional appearance
The cross-section of a primary stem may have a more or less angular to cir-cular outline. However, it can take on one of a wide range of forms, some of which assist in the identification of a family, as in Labiateae, where the sec-tion is square, or may help to distinguish genera; for example many Carex species have stems with a triangular cross-section. Often the outline is modified near to nodes or in regions of leaf insertion. Sometimes a wing or ridge of tissue in line with the sides of a petiole may continue down the internode as in, for example, Lathyrus. In general, the outline of the section taken in the middle of an internode would be described for comparative purposes.
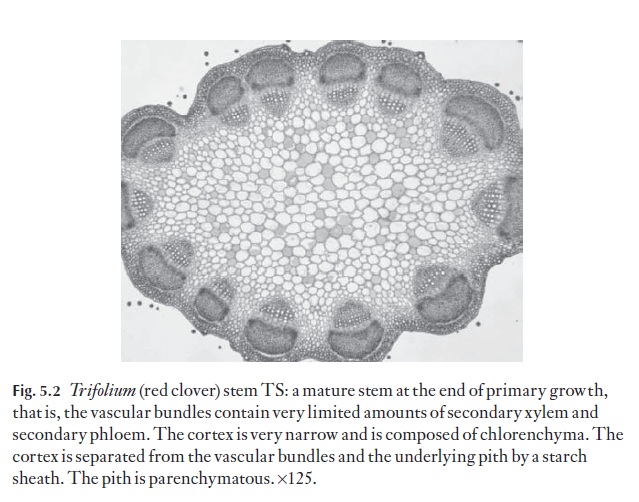
As indicated in the introduction, stems have all or most of the following tissues, working from the outside inwards: epidermis, hypodermis, cortex (with both collenchyma and chlorenchyma, or either alone), an endo-dermoid layer (or a well-defined starch sheath), vascular bundles in one or more rings, or apparently scattered, and a central ground tissue or pith (Fig. 5.2).
Sometimes a pericycle can be distinguished, but this is normally regarded as part of the phloem. A true endodermis with Casparian strips is rarely present.
Epidermis
The epidermis can be one or more layered. The cells may be similar in form to those of the leaf of the same species, but more often they show more elon-gation in the direction parallel with the stem axis, and their anticlinal walls as seen in surface view are often not markedly sinuous. The outer wall is usually thicker than the anticlinal or inner walls. The proportion of cell length to cell width, or height to width as seen in TS, can be used with cau-tion as a diagnostic feature, but actual measurements are not normally suf-ficiently constant to be used in identification or classification. A range of the normal measurements of a cell, with an average figure, should be quoted in diagnostic descriptions where a sufficiently large sample has been exam-ined in order to obtain sound data.
Cuticular sculpturing, fine surface features which include very small pa-pillae (micropapillae) and fine ridges (striae) in a variety of arrangements, may be similar to that in the leaves of the same species, but frequently dif-fers in details.
Hypodermis
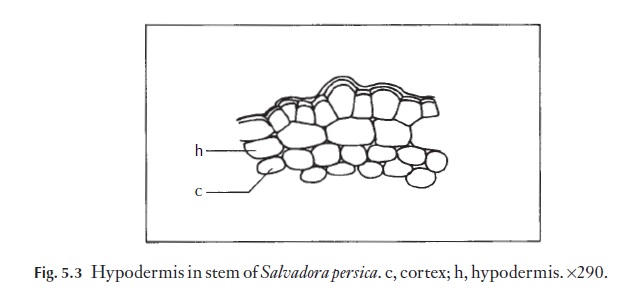
The layer of cells under the epidermis is called the hypodermis. This may show distinct differences from the next cortical layer, but in many taxa is defined only by its position. When it is not different in appearance it is com-monly described by systematic plant anatomists as being absent. A distinct hypodermis is found in Salvadora persica, for example Fig. 5.3. However, be-cause of the sporadic occurrence of the distinct hypodermis in the taxa of vascular plants, its presence or absence is of little taxonomic and small diag-nostic value (except at the species level).
Stomata
In taxa with well-developed leaves the stem stomata tend to be much more sparse, but of the same type as in the leaf of the same species. When the stem is a principal photosynthetic organ, either supplementing or replacing leaves, stomata tend to occur in a higher frequency. Often the guard cells are aligned parallel to the long axis of the stem and the stomata appear in rows.
Trichomes
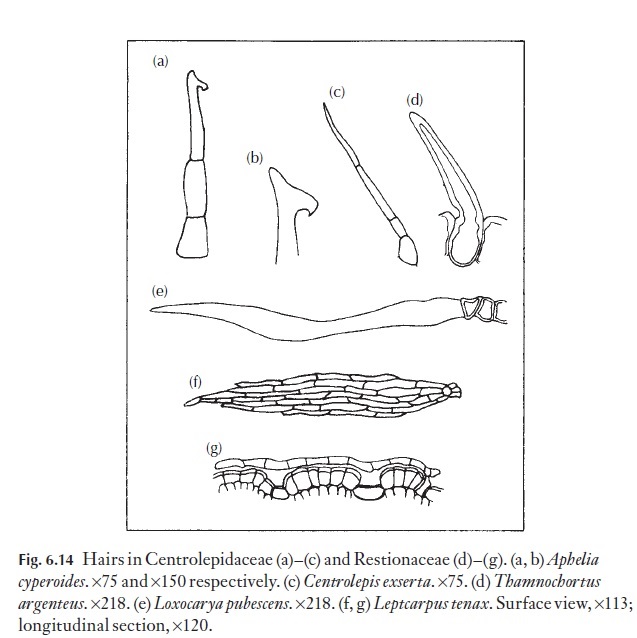
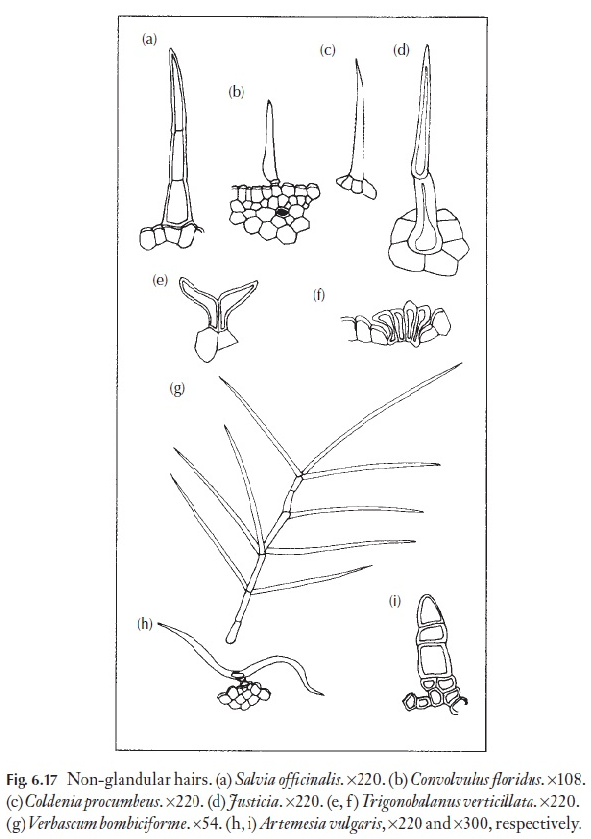
Hairs, papillae and scales exhibit the same wide range as on the leaf, and ex-amples are shown in Figs 6.14–6.17. The type of hair can be of diagnostic value at species level, sometimes also at genus level, but rarely at family level.
Silica bodies
Silica bodies are often present in the stem epidermis or in other parts of the stem in species which have them in the leaves. There are also leafless species of some families which have silica bodies in the stems for example Lepyrodia scariosa, Restionaceae. In the epidermis, the cells most likely to have silicabodies are those above fibre strands or girders. The range of form is shown in Fig. 6.24.
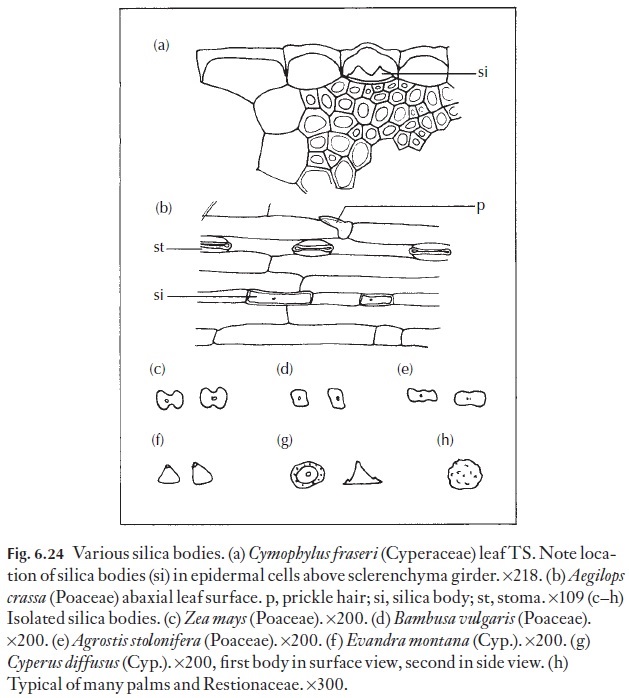
![]()
![]()
Cortex
The cortex can be very narrow, and composed of few cell layers, or wide and multilayered. The cortical zone is traditionally regarded as extending from epidermis or hypodermis to an inner boundary inside which vascular bun-dles are present. The inner boundary is often very indistinct, and some-times vascular bundles from leaf traces may be present among cells which clearly belong to the cortex itself. Again we have an example of a tissue which is difficult to define clearly.
Chloroplasts may be present in the collenchyma cells of the outer cortex, or in more or less well-defined layers of parenchymatous cells, as well as in palisade cells or cells of various shapes. Stems of leafless plants, for example some Juncus species, often have a very formal and regular chlorenchyma ar-rangement. There are a few herbaceous plants which lack chloroplasts in cortical tissues, and these plants normally figure among those with abnor-mal modes of nutrition, for example Orobanche.
In species with a wide cortex, the cells of inner layers are normally larger than those of outer layers and have few, if any, chloroplasts. In aquatic plants, large, formal air spaces may be present in the cortex which itself merges with central parenchymatous tissues.
Fibres and sclereids are a prominent feature in the cortex of many species. Often the grouping of fibres into strands with well-defined cross-sectional outlines and in characteristic positions in the cortex will help in the identi-fication of a plant. Fibres can show individual peculiarities, which enable one to identify even isolated strands. This is particularly true for fibres which are of economic importance, many of which are cortical in origin, for example Linum, flax, and Boehmeria.
Crystals and tannin often occur in cells of the cortex and the central ground tissue. Cluster crystals or druses are probably the most common type, but solitary crystals of various shapes and sizes, similar to the range exhibited in leaves, are of widespread occurrence. Raphides are not of wide-spread occurrence in dicotyledons.
Endodermis
In some plants the inner boundary of the cortex is well differentiated into an endodermis and shows Casparian strips, for example Hydrocharita-ceae. In Helianthus, the cells are well-defined and are rich in stored starch but lack Casparian strips, thus they constitute a ‘starch sheath’. In many other plants where the cells in this zone are morphologically distinctive but ![]()
![]() lack Casparian strips or stored starch, they are probably best called an ‘endodermoid layer’. Some people prefer to use the term endodermis for this cell layer even when Casparian strips cannot be seen.
lack Casparian strips or stored starch, they are probably best called an ‘endodermoid layer’. Some people prefer to use the term endodermis for this cell layer even when Casparian strips cannot be seen.
Vascular and strengthening tissue
Many monocotyledons, but few dicotyledons, have a well-developed cylin-der of sclerenchyma to the inner side of the endodermoid layer. In it are embedded some of the vascular bundles, often all the small bundles, and sometimes some of the larger bundles as well. Most dicotyledons with secondary growth in thickness lack such a cylinder. Their ‘open’ vascular bundles, each with a cambium, are arranged in a ring. Each bundle may have a cap of fibres to its outer side, but the flanks of the bundles are not en-closed, so enabling unhindered development of the interfascicular cam-bium to produce a continuous cambial cylinder in secondary growth.
In many of those monocotyledons lacking a sclerenchymatous cylinder, individual bundles have a sheath of sclerenchyma. This is frequently only a few layers thick on the flanks, and several layers thick at the xylem and phlo-em poles. An outer parenchymatous sheath features in a number of these plants. Because there is no cambium as such, the enclosing of vascular tis-sues in these plants does not affect normal growth and development. In the monocotyledons which have a vascular plexus at the nodes, the sclerenchy-ma sheaths become discontinuous. If there is an intercalary meristem, the bundle sheaths are poorly developed in the meristematic zone.
In monocotyledonous stems the vascular bundles are either collateral, with one xylem and one phloem pole, or amphivasal, with the xylem encir-cling the phloem. Amphivasal bundles are frequent in rhizomes (which are modified stems) and less frequent in inflorescence axes (flower stems). Figure 5.4 shows a range of bundle types. Dicotyledons usually have open bundles, but climbers, for example Cucurbitaceae (cucurbits), may not develop interfascicular cambia, and the bundles remain as discrete strands. Since such bundles are present in a compressible parenchymatous matrix, twisting and distorting of such stems during the process of climbing does little damage to the vascular bundles themselves. In cucurbits and a number of other climbers the phloem is particularly well developed. It is present in two strands, one on either side of the xylem on a radial axis as seen in trans-verse section. Bundles of this kind are called bi-collateral. Because they occur in relatively few families their presence in a sample to be named be-comes most helpful in reducing the field for further analysis.
Vascular bundles, which have phloem surrounding the xylem, are termed amphicribal.
There are plants with anomalous growth in which individual bundles have cambium and extend radially in secondary growth, without uniting laterally, as for example in the Piperaceae (Fig. 5.4). Several other abnormal forms of bundle arrangement are found among the dicotyledons.
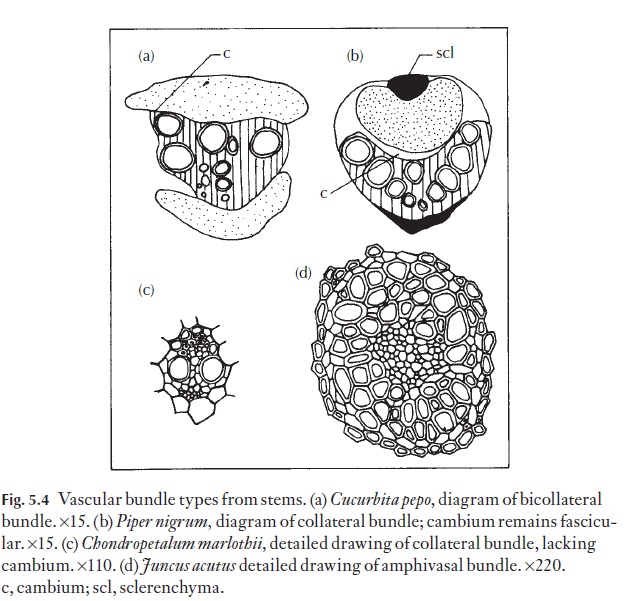
It is a dangerous practice to attempt to define various types of vascular bundles on the observations made from a few transverse sections of mono-cotyledon stems. This is because over its length, a single bundle can exhibit changes in its cross-sectional appearance. A newly entering leaf trace can look different from a main axial bundle but the two are parts of the same strand, the first being the increment before bridging and connecting with the strands from other parts of the plant.
Related Topics