Chapter: Essential Microbiology: Microbiology: What, Why and How?
How do we know? Microbiology in perspective: to the ‘golden age’ and beyond
How do we
know? Microbiology in perspective: to the ‘golden age’ and beyond
We have learnt an astonishing amount about the
invisible world of microorganisms, particularly over the last century and a
half. How has this happened? The penetrating insights of brilliant individuals
are rightly celebrated, but a great many ‘breakthroughs’ or ‘discoveries’ have
only been made possible thanks to some (frequently unsung) development in
microbiological methodology. For example, on the basis that ‘seeing is
believing’, it was only when we had the means to see microorganisms under a micro-scope that we could prove their
existence.
Microorganisms had been on the Earth for some 4000
million years, when Antoni van Leeuwenhoek started out on his pioneering
microscope work in 1673. Leeuwen-hoek was an amateur scientist who spent much
of his spare time grinding glass lenses
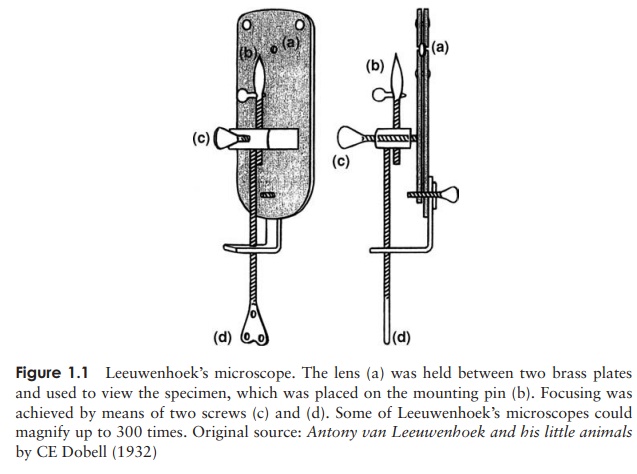
to produce simple microscopes (Figure 1.1). His
detailed drawings make it clear that the ‘animalcules’ he observed from a
variety of sources included representatives of what later became known as
protozoa, bacteria and fungi. Where did these creatures come from? Arguments
about the origin of living things revolved around the long held belief in
spontaneous generation, the idea that living organisms could arise from
non-living matter. In an elegant experiment, the Italian Francesco Redi
(1626–1697) showed that the larvae found on putrefying meat arose from eggs
deposited by flies, and not spontaneously as a result of the decay process.
This can be seen as the beginning of the end for the spontaneous generation
theory, but many still clung to the idea, claiming that while it may not have
been true for larger organisms, it must surely be so for minute creatures such
as those demonstrated by Leeuwenhoek. Despite mounting evidence against the
theory, as late as 1859, fresh ‘proof’ was still being brought forward in its
support. Enter onto the scene Louis Pasteur (1822–1895), still arguably the
most famous figure in the history of microbiology. Pasteur trained as a
chemist, and made a lasting contribution to the science of stereochemistry
before turning his attention to spoilage problems in the wine industry. He
noticed that when lactic acid was produced in wine instead of alcohol,
rod-shaped bacteria were always present, as well as the expected yeast cells.
This led him to believe that while theyeast produced the alcohol, the bacteria
were responsible for the spoilage, and that both types of organism had
originated in the en-vironment. Exasperated by continued efforts to
substantiate the theory of spontaneous generation, he set out to disprove it
once and for all. In response to a call from the French Academy of Science, he
carried out a series of experiments that led to the ac-ceptance of biogenesis, the idea that life arises
only from already existing life. Using his famous swan-necked flasks (Figure
1.2), he demonstrated in 1861 that as long as dust
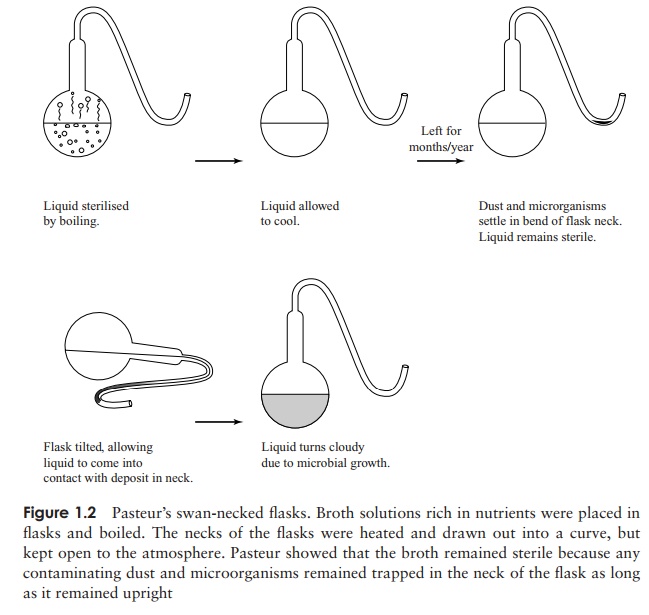
particles (and the microorganisms carried on them)
were excluded, the contents would remain sterile. This also disproved the idea
held by many that there was some element in the air itself that was capable of initiating
microbial growth. In Pasteur’s words ‘. . . . the doctrine of spontaneous
generation will never recover from this mortal blow. Thereis no known circumstance in which it can be affirmed that
microscopic beings came into the world without germs, without parents similar
to themselves.’ Pasteur’s findingson wine contamination led inevitably to
the idea that microorganisms may be also be responsible for diseases in humans,
animals and plants.
The notion that some invisible (and therefore, presumably,
extremely small) living creatures were responsible for certain diseases was not
a new one. Long before micro-organisms had been shown to exist, the Roman
philosopher Lucretius (∼98–55 BC) and much later the
physician Girolamo Fracastoro (1478–1553) had supported the idea. Fracastoro
wrote ‘Contagion is an infection that passes from one thing to another’ and
recognised three forms of transmission: by direct contact, through inanimate
objects and via the air. We still class transmissibility of infectious disease
in much the same way today. The prevailing belief at the time, however, was
that an infectious disease was due to something called a miasma, a poisonous vapour arising from dead or diseased bodies, or
to an imbalance between the four humours of the body (blood, phlegm, yellow
bile and black bile). During the 19th century, many diseases were shown, one by
one, to be caused by microorganisms. In 1835, Agostino Bassi showed that a
disease of silkworms was due to a fungal infection, and 10 years later, Miles
Berkeley demonstrated that a fun-gus was also responsible for the great Irish
potato blight. Joseph Lister’s pioneering work on antiseptic surgery provided
strong, albeit indirect, evidence of the involvement of mi-croorganisms in
infections of humans. The use of heat-treated instruments and of phenol both on
dressings and actually sprayed in a mist over the surgical area, was found
greatly to reduce the number of fatalities following surgery. Around the same
time, in the 1860s, the indefatigable Pasteur had shown that a parasitic
protozoan was the cause of another disease of silkworms called pebrine´, which had devastated the
French silk industry.
The first proof of the involvement of bacteria in disease and
the definitive proof of the germ theory of disease came from the German Robert
Koch. In 1876 Koch showedthe relationship between the cattle disease anthrax and a bacillus which we now know
as Bacillus anthracis. Koch infected
healthy mice with blood from diseased cattle and sheep, and noted that the
symptoms of the disease appeared in the mice, and that rod shaped bacteria
couldbe isolated from their blood. These could be grown in culture, where they
multiplied and produced spores. Injection of healthy mice with these spores (or
more bacilli) led them too to develop anthrax and once again the bacteria were
isolated from their blood. These results led Koch to formalise the criteria
necessary to prove a causal relationship between a specific disease condition
and a particular microorganism. These criteria became known as Koch’s postulates, and are still in use
today.
Despite their value, it is now realised that Koch’s
pos-tulates do have certain limitations. It is known for ex-ample that certain
agents responsible for causing disease (e.g. viruses, prions:) can’t be grown invitro, but only in host cells. Also,
the healthy animalin Postulate 3 is seldom human, so a degree of extrapo-lation
is necessary – if agent X does not cause disease in
a laboratory animal, can we be sure it won’t in
humans? Furthermore, some diseases are caused by more than one organism, and
some organisms are responsible for more than one disease. On the other hand,
the value of Koch’s postulates goes beyond just defining the causative agent of
a particular disease, and allows us to ascribe a specific effect (of whatever
kind) to a given microorganism.
Critical to the development of Koch’s postulates was
the advance in culturing techniques, enabling the isola-tion and pure culture
of specific microorganisms. The development of pure cultures revolutionised
microbiology, and within the next 30 years or so, the pathogens responsible for
the majority of common human bacterial diseases had been isolated and
identified. Not without just cause is this period known as the ‘golden age’ of
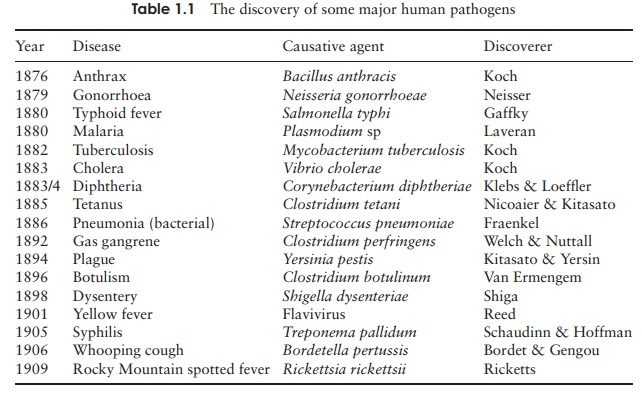
microbiology! Table 1.summarises the discovery of some major human pathogens.
Koch’s greatest achievement was in using the
ad-vances in methodology and the principles of his own postulates to
demonstrate the identity of the causative agent of tuberculosis, which at the
time was responsible for around one in every seven human deaths in Europe.
Although it was believed by many to have a microbial
cause, the causative agent had never been observed, either in culture or in the
affected tissues. We now know that Mycobacterium
tuberculosis (the tubercle bacillus) is very difficult to stain by
conven-tional methods due to the high lipid content of the cell wall surface.
Koch developed a staining technique that enabled it to be seen, but realised
that in order to satisfy his own postulates, he must isolate the organism and
grow it in culture. Again, there were technical difficulties, since even under
favourable conditions, M. tuberculosis
grows slowly, but eventually Koch was able to demonstrate the infectivity of
the cultured organisms towards guinea pigs. He was then able to isolate them
again from the dis-eased animal and use them to cause disease in uninfected
animals, thus satisfying the remainder of his postulates.
Although most bacterial diseases of humans and their
aetiological agents have now been identified, important variants continue to
evolve and emerge. Notable exam-ples in recent times include Legionnaires’
disease, an acute respiratory infection caused by the previously unrecognised
genus, Legionella, and Lyme disease,
a tickborne infection first described in Connecticut, USA in the mid-1970s.
Also, a newly recognised pathogen, Helicobacter
pylori, has been shown to play an important (and previously unsuspected)
role in the development of peptic ulcers. There still remain a few diseases
that some investigators suspect are caused by bacteria, but for which no
pathogen has been identified.
Following the discovery of viruses during the last decade
of the 19th century, it was soon established that many diseases of plants,
animals and humans were caused by these minute, non-cellular agents.
The major achievement of the first half of the 20th
century was the development of antibiotics and other antimicrobial agents.
Infectious diseases that previously accounted for millions of deaths became
treatable by a simple course of therapy, at least in the affluent West, where
such medications werereadily available.
If the decades either side of 1900 have become known
as the golden age of microbiology, the second half of the twentieth century
will surely be remembered as the golden age of molecular genetics. Following on
from the achievements of others such as Griffith and Avery, the publication of
Watson and Crick’s structure for DNA in 1953 heralded an extraordinary 50 years
of achievement in this area, culminating at the turn of the 21st century in the
completion of the Human Genome Project.
What, you might ask, has this genetic revolution to
do with microbiology? Well, all the early work in molec-ular genetics was
carried out on bacteria and viruses and microbial systems have also been
absolutely central to the development of genetic engineering over the last
three decades. Also, as part of the Human Genome Project, the genomes of
several microorganisms have been decoded, and it will become increasingly easy
to do the same for others in the future, thanks to methodological advances made
during the project. Having this information will help us to understand in
greater detail the disease strategies of microorganisms, and to devise ways of
countering them.
As we have seen, a recurring theme in the history of
microbiology has been the way that advances in knowledge have followed on from
methodological or technological developments, and we shall refer to a number of
such developments during the course of this book. To conclude this introduction
to microbiology, we shall return to the instrument that, in some respects,
started it all. In any microbiology course, you are sure to spend some time
looking down a microscope, and to get the most out of the instrument it is
essential that you understand the principles of how it works. The following
pages attempt to explain these principles.
Related Topics