Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: Teleosts at last I: bonytongues through anglerfishes
Neognathi
Neognathi
The remaining Euteleosts, all 336 families and 17,000+species, are collectively placed in the unranked taxonomiccategory Neognathi, which consists of the order Esociformes(10 species) and the Neoteleostei, the latter also anunranked taxonomic level. The esociform pikes and mudminnowsare viewed as the sister group of all higher euteleostsbecause they share a distinct type of tooth attachmentand a skeleton made up of acellular bone (Johnson &Patterson 1996). Consensus on this arrangement is by nomeans universal, with some authors contending that esociformsare protacanthopterygians and a sister taxon to salmoniforms(see Nelson 2006 for a review).
Order Esociformes
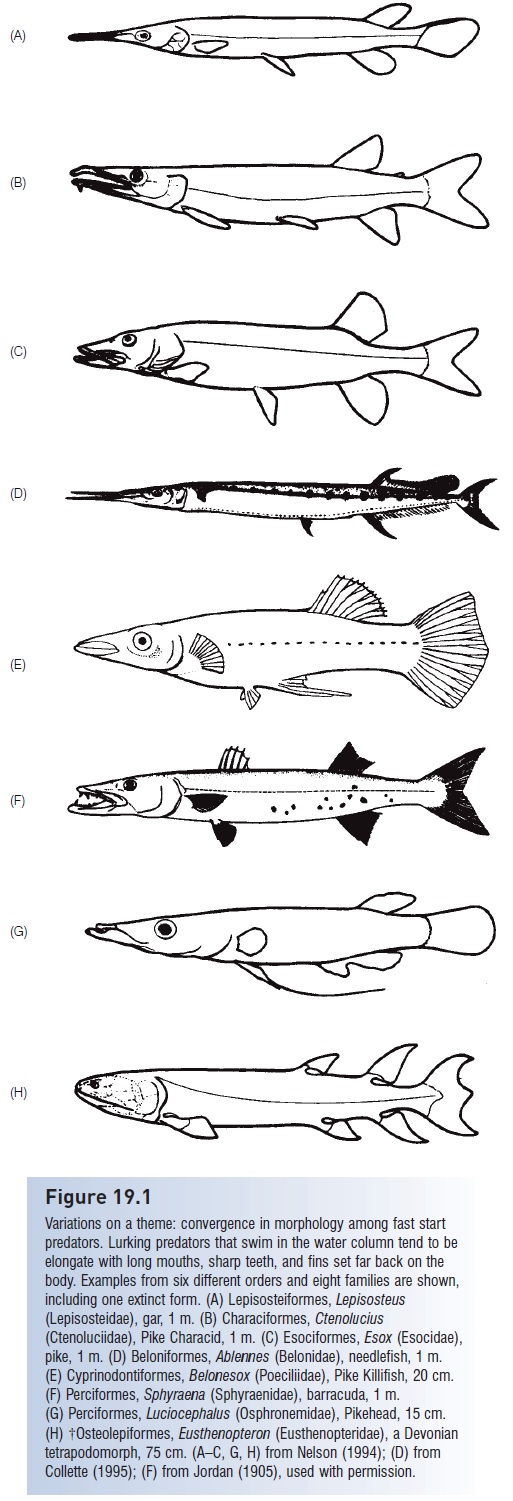
The esociforms consist of two temperate families of freshwaterfishes in which the maxillary bone is included in thegape but is toothless, and in which the median fins arelocated relatively far back on the body. Esocids include theGrass, Redfin, and Chain pickerels, the Northern and Amurpikes, and the superpredatory 1.4 m Muskellunge,Esoxmasquinongy (Fig. 14.19). The Northern Pike, E. lucius,has the most widespread natural east–west distributionof any completely freshwater fish, occurring across thenorthern portions of North America, Europe, and Asia(=circumpolar distribution). The family goes back into theCretaceous, and one Paleocene species is very similar to theNorthern Pike despite a 62-million-year separation. Anatomicaland behavioral convergences between pikes andbarracudas, the latter a higher acanthopterygian, are fairlyremarkable (see Pursuit and Fig. 19.1). Themudminnows and blackfishes (Umbridae) are also scatteredacross northern North America, Europe, and Asia. TheCentral Mudminnow, Umbra limi, has a remarkable abilityto survive in high-latitude lakes that become ice-coveredand oxygen-poor through much of the winter (see Seasonal cycles).
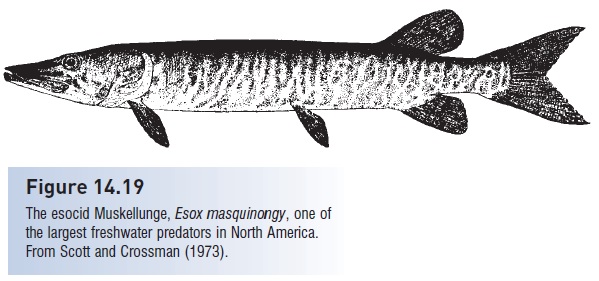
Figure 14.19
The esocid Muskellunge, Esox masquinongy, one ofthe largest freshwater predators in North America.From Scott and Crossman (1973).
Neoteleostei
All of the fishes above the esociforms are considered to beneoteleosts, a category without formal rank that lies somewherebetween a subdivision and a superorder (Fig. 14.20).Eight neoteleostean superorders are recognized, includingsix relatively specialized deepsea and pelagic superorders and twovery diverse, advanced superorders, the paracanthopterygiansand acanthopterygians. Neoteleosts as a taxon areconsidered to be monophyletic on the basis of four skulland jaw characters possessed by the different members thatare lacking in more primitive fishes:
1 The manner in which the vertebral column connects to the back of the skull changes. In the neoteleosts, thefirst vertebra articulates with three bones of the skull(basioccipital and the two exoccipitals), whereas inmore primitive teleosts it articulates with only theunpaired basioccipital.
2 A muscle, the retractor dorsalis, connects the vertebralcolumn with dorsal elements of the upper pharyngealjaws and pulls those jaws posteriorly.
3 A shift occurs in the insertion position ofanother muscle, one of the internal levators thatoriginates on the base of the skull and lifts thepharyngeal jaws.
4 A unique hinged manner of attaching teeth to the jawsdevelops, allowing the tooth to be depressed towardsthe back of the mouth.
In addition, a trend toward more anteriorly located pelvicsand more laterally located pectorals is evident during neoteleosteanphylogeny, and acellular bone (skeletal materiallacking bone cells) occurs in most neoteleosts, whereas
more primitive groups have bone cells (Smith 1988).
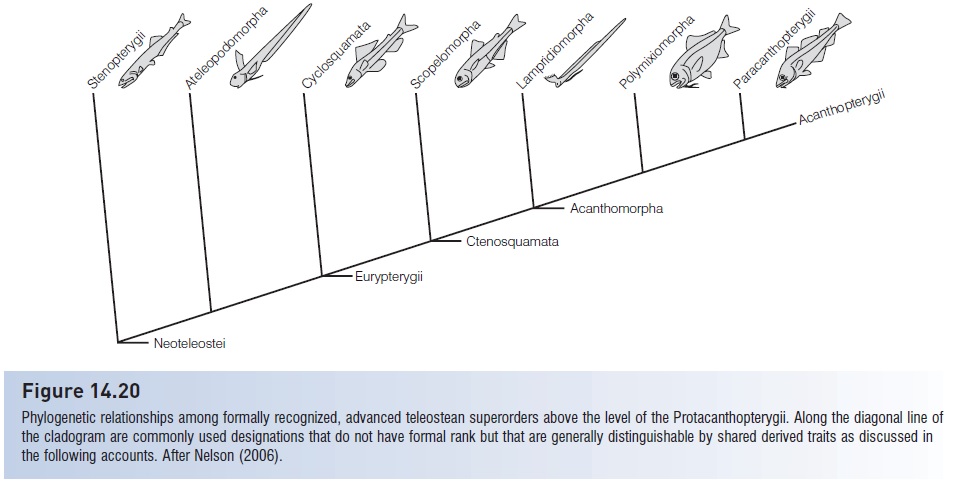
Figure 14.20
Phylogenetic relationships among formally recognized, advanced teleostean superorders above the level of the Protacanthopterygii. Along the diagonal line ofthe cladogram are commonly used designations that do not have formal rank but that are generally distinguishable by shared derived traits as discussed inthe following accounts. After Nelson (2006).
Superorder Stenopterygii
Stomiiforms are all deepsea fishes of the mesopelagic andbathypelagic regions (open water, between 200 and about4000 m depths) and are often characterized by long teethuniquely attached to jaw bones, large mouths, histologicallyunique photophores (light organs) that include a duct, anda peculiar ventral adipose fin ahead of the anal fin in some.Gonostomatid bristlemouths (Cyclothone) and phosichthyidlightfishes (Vinciguerria) may be the most abundantand widely distributed vertebrates on earth. The sternoptychidmarine hatchetfishes possess several structural specializationsthat emphasize the vertical plane and bodycompression. The mouth opens vertically, the photophorespoint down, a preopercular spine points upwards, thepelvic bones are oriented vertically, the lateral compressionof the body is heightened by an abdominal keel-like structure,and a bladelike structure preceding the dorsal fin ismade up of dorsal pterygiophores that project through theback of the fish. Pterygiophores normally serve as the basalsupport for median fins, not as elements of a fin itself. Idiacanthineblack dragonfishes have a larval form with eyesat the ends of elongated stalks. Stomiiforms show manytraits common to other, unrelated deepsea fishes that aregenerally viewed as convergent adaptations to the light- andfood-limited conditions of the deep sea (see The deep sea).
Superorder Ateleopodomorpha
The ateleopodiform jellynose fishes are an unusual groupof bulbous-headed, elongate species that swim just abovethe bottom in deep water. Their skeleton is largely cartilaginous.Their large and pointed head, exaggerated analfin, and relatively pointed tail are all traits they share withother deep, benthopelagic fishes such as chimaeras, spinyeels, halosaurs, eucla cods, rattails, and grenadiers.
Superorder Cyclosquamata
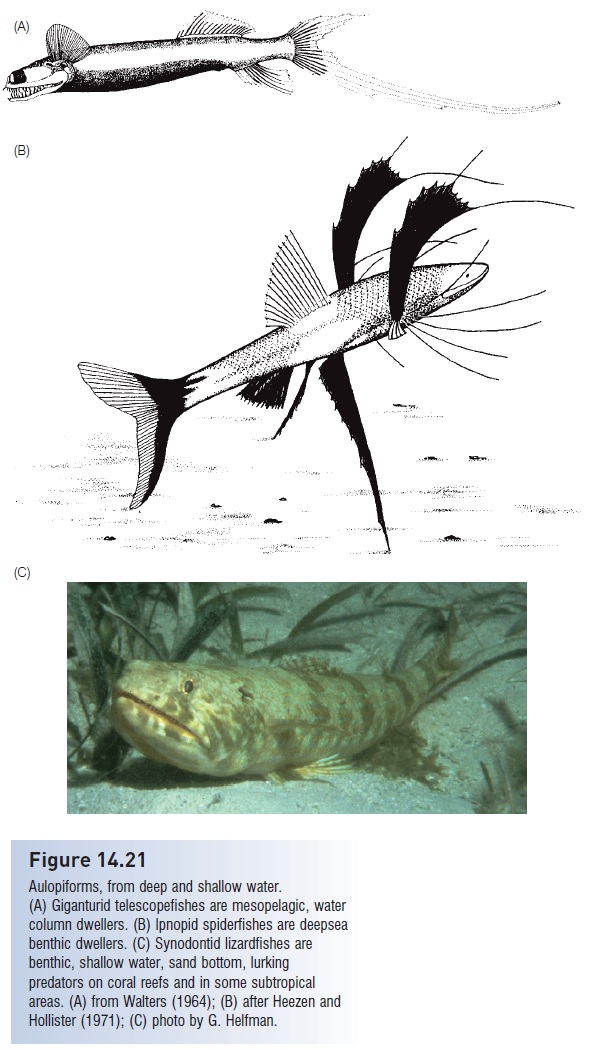
Fishes more advanced than ateleopodomorphs are sometimesreferred to as eurypterygians (“wide or broad fins”).The most primitive superorder in this group is the Cyclosquamata(“cycloid scales”), which contains one large orderof almost entirely deepsea fishes. Aulopiforms have a surprisinglyextensive fossil record (approximately six familiesand perhaps 20 genera) for a group that today is largelyopen water and deep sea. The extant deepsea groups includethe truly bizarre giganturid telescope fishes (Fig. 14.21A)which undergo a spectacular metamorphosis during whichthe premaxillary, palatine, orbitosphenoid, parietal, symplectic,posttemporal, supratemporal, hyoid, and cleithralbones in the head region are lost, as are several gill arches,the gill rakers, and the gas bladder. The larvae possess anadipose fin, pelvic fin, and branchiostegal rays that are alsolost during metamorphosis. The adults possess large tubulareyes, a huge mouth, fl exible teeth, an expandable stomach,pectoral fins located exceptionally high on the body abovethe eyes, loose skin, and a peculiar tail with the ventral lobeextending far beyond the dorsal lobe.
Chlorophthalmid greeneyes are the firstfishes encounteredin this survey that are hermaphroditic, as are members of other families in this order. All of the more primitivegroups so far have been distinctly gonochoristic, whichmeans that an individual is only one sex throughout its life.Hermaphroditism of some form is surprisingly common inhigher teleosts. This dichotomy between primitive andadvanced teleosts suggests that sexual lability – either interms of initial sexual determination or of an ability tochange sex later in life – represents a derived trait thatbecame differentially retained or independently evolvedafter the neoteleostean or eurypterygian level of developmentwas reached.
Included among the ipnopids are the often illustratedspiderfishes or tripodfishes, which have greatly elongatedpectoral, pelvic, and caudal rays; they use the pelvics andcaudal to form a tripod for resting on sediments of the deepocean floor (Fig. 14.21.B). The best known shallow representativesof this order are the synodontid lizardfishes,which are common benthic inhabitants of coral reefs worldwide(and whose name should not be confused with thesynodid upsidedown catfishes) (Fig. 14.21C). Lizardfishesare closely related to the secondarily pelagic Bombay ducks(Harpadon) that support an important fishery in the IndianOcean. The order includes the alepisaurid lancetfishes,which are large (up to 2 m) mesopelagic predators on otherfishes. Lancetfishes are distinguished by their large, sail-likedorsal fin that extends from the head almost to the caudalpeduncle (function unknown). These fishes, which lookvery much like scombroid snakemackerels, have proven agreat boon to deepsea taxonomists as several species ofmesopelagic fishes have been described from the stomachcontents of alepisaurids (see The deep sea).
Superorder Scopelomorpha
All fishes above the level of the Cyclosquamata have lostthe fi fth pharyngeal toothplate and the muscle that lifts itJohnson 1992). These advanced groups are often termedthe Ctenosquamata in reference to the predominance ofctenoid scales among them. One order of deepsea fishes,the myctophiforms, makes up the superorder Scopelomorpha(Fig. 14.22). Myctophiforms have lost the fi fthpharyngeal toothplate but most still have cycloid scales,justifying their primitive status among ctenosquamates.Their relative primitiveness is also shown in their retentionof an adipose fin, but they are advanced in that the maxillais excluded from the gape. The myctophid lanternfishes,with about 240 species, are an important group of mesopelagicdeepsea fishes in terms of diversity, distribution, andnumbers of individuals. They occur in all seas, from the Arctic to the Antarctic, and are the prey of numerous otherfishes as well as of marine mammals. They make up a largefraction of the deep scattering layer – a diverse assemblageof fishes and invertebrates that live at mesopelagic depths

Figure 14.22
A myctophid lanternfish, Diaphus mollis, about4 cm long. The round structures along the ventralhalf of the body are light-emitting photophores.From Nafpaktitis et al. (1977), used withpermission.
Related Topics