Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: Teleosts at last I: bonytongues through anglerfishes
Acanthomorpha: thespiny teleosts
Acanthomorpha: thespiny teleosts
Neoteleosts above scopelomorphs possess true fin spinesand are termed acanthomorphs. The appearance of true finspines, rather than hardened segmented rays, marks a majorevolutionary step in the evolution of bony fishes. Truespines occur in the dorsal, anal, and pelvic fins of higherteleosts. Spines develop when the two halves of the primitivelypaired and jointed dermal fin rays fuse into a single,unsegmented structure. Several other characteristics, mostlyassociated with improved locomotion and feeding, alsomark the ascendancy of teleosts above the ctenosquamatelevel of development (see Division Teleostei).Locomotion was improved by the strengthening of vertebralaccessories (zygapophyses), providing body stiffeningand better attachment for muscles. These changes alloweda shift from slow, sinusoidal motion of the entire body torapid oscillation of the tail region, driven by tendonsattached to the tail base (see Locomotion: movementand shape). The tail itself also underwent considerablemodification. Pharyngeal teeth diversified and the maxillashifted from a tooth-bearing bone to a structure that helpspivot the premaxilla, making mouth protrusion and suctionmore effective. These advances probably made possible theexplosive radiation of spiny-rayed teleosts, known collectivelyas acanthomorphs, during the Early Cenozoic.The acanthomorphs include the superorders Lampriomorpha,Polymixiomorpha, Paracanthopterygii, and Acanthopterygii.
Superorder Lampriomorpha
As is often the case, the most primitive members of a majortaxon retain certain ancestral traits but possess others indicativeof advanced status. Lampriforms lack true spines, butthe maxilla helps move the premaxilla and bears no teeth.The connection between the two upper jaw bones and themanner in which they slide to protrude the mouth areunique, so much so that the group used to be referred toas the Allotriognathi, or “strange jaws”. The seven familiesof lampriforms are almost all open water oceanic fisheswith unusual body and fin proportions. Opahs are relativelylarge (up to 1.8 m, 70 kg), oval-shaped, colorful pelagicpredators on squids and other fishes (Fig. 14.23A). The30 cm long tube-eye (Stylephorus) is capable of an almost40-fold enlargement of its mouth volume during feeding,which is probably a record among vertebrates (see Jaw protrusion: the great leap forward). The remainingfive families contain mostly large and rare, elongate, pelagicfishes with long dorsal fins. Crestfishes and radiicephalidshave ink sacs that they discharge through their cloaca. In trachipterid ribbonfishes, the caudal fin is made up only ofthe upper lobe, which sits inclined at a right angle upward.The oarfish, Regalecus, reaches a confi rmed length of over7 m and perhaps as much as 11 m, making it the longestextant teleost; uncertainty arises because intact specimensare seldom found (Fig. 14.23B). Despite their size, they areapparently planktivores that hover head up in the watercolumn, swimming via rapid undulations of the long dorsalfin. Their name comes from bladelike expansions at the endof the pelvic fins. Specimens are uncommon, many obtainedafter storms when the bodies of these strange-looking fishesare tossed up on beaches. The oarfish, with its bluish-silverybody, scarlet dorsal crest of elongate fin rays, and deep redfins, is likely responsible for many sea serpent sightings,particularly those referring to monsters “having the headof a horse with a fl aming red mane” (Norman & Fraser1949, p. 113).
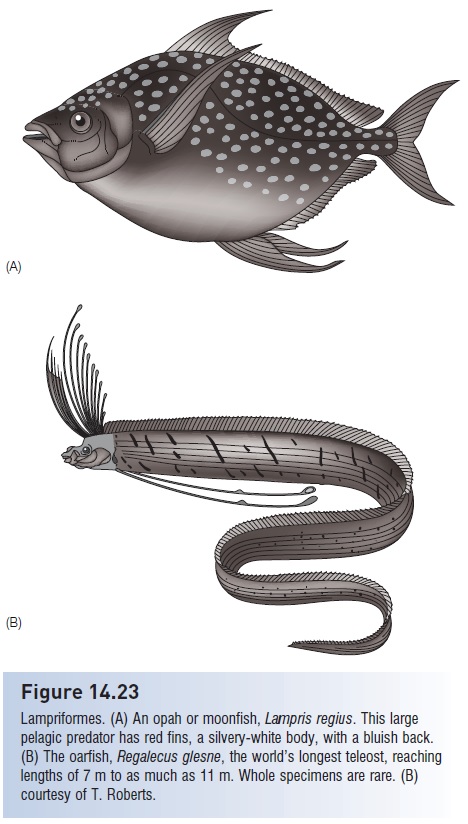
Figure 14.23
Lampriformes. (A) An opah or moonfish, Lampris regius. This largepelagic predator has red fins, a silvery-white body, with a bluish back.(B) The oarfish, Regalecus glesne, the world’s longest teleost, reachinglengths of 7 m to as much as 11 m. Whole specimens are rare. (B)courtesy of T. Roberts..
Superorder Polymixiomorpha
The taxonomic status of this enigmatic family has been thesubject of considerable debate (see Johnson & Patterson1993a, 1993b; Nelson 1994, 2006). Beardfishes possessadvanced characters such as four to six true spines in thedorsal fin and four spines in the anal fin, and their pelvicfins are located fairly forward on the body. Yet they retaintwo sets of intermuscular bones, the epineurals and epipleurals,a characteristic of more primitive taxa, and have aunique arrangement of jaw muscle ligaments. This combinationof primitive, advanced, and unique characteristicscomplicates determination of their relationships. At differenttimes, they have been classified as a sister taxon to theremaining acanthomorphs, with the primitive paracanthopterygianpercopsiforms (trout-perches and relatives), or withprimitive acanthopterygian beryciforms (squirrelfishes andrelatives). We follow Nelson (2006) and place them betweenlampriomorphs and paracanthopterygians, but with minimalcertainty. Beardfishes have large eyes, chin barbels, are about30 cm long, and live at moderate depths (200–600 m).
The 36 families of recognized paracanthopterygians representa major and contentious side branch during the evolutionof advanced acanthomorphs. They are weakly definedby a number of characters, chiefl y involving the caudalskeleton and holes in the skull through which cranial nervespass (Nelson 1994), but “. . . there is no fi rm basis to believe[the superorder] is monophyletic” (Nelson 2006, p. 232).Their ecological success is largely as benthic, marine fishesthat are nocturnally active or live in permanently darkenedwaters, such as the bathypelagic region of the deep sea orin caves. Only about 20 relatively primitive paracanthopterygians,out of a total 1340 species, live in fresh water.Manyparacanthopterygians have sonic muscles on their gasbladders and produce sounds.
At the base of the paracanthoptgerygians are the percopsiforms,small (<20 cm), freshwater fishes, eight of nine ofwhich live in eastern North America. The Trout-perch,Percopsis omiscomaycus, and its Columbia River congener,P. transmontana, are the most advanced fishes we willencounter with an adipose fin. This seeming primitive conditionis interesting given that modern percopsiformspossess several traits suggesting a reversal of the evolutionarytrends of advanced teleosts, including fossil percopsiforms.The modern species have fewer fin spines, morevertebrae, and a more posteriorly located pelvic girdle thanoccurred in fossil forms. The aphrododerid Pirate Perch,Aphrododerus sayanus, is a swamp dweller with the distinctionof having its anus move from just anterior of the analfin in juveniles to the throat region of adults. Speculationabounded on the functional reasons for this anatomicalanomaly. Recent observations reveal that Pirate Perch arenocturnally active fish that hide and spawn in dense vegetationand root mats (Fletcher et al. 2004; Tiemann 2004).Females push head first into such root mats to deposit eggs,and males enter the same spot shortly thereafter to depositsperm; hence the anterior position of the anus allows depositionof eggs and sperm deep into a protected, otherwiseinaccessible area (Fig. 14.24). By day, fish emerge fromvegetation refuges only far enough to expose the head andanus, at which time they defecate. The jugular position ofthe anus would permit defecation without altering waterquality in the refuge and would also minimize exposure topredators. As is so often the case, seemingly specializedtraits often confer multiple functional benefits.
The amblyopsid cavefishes include six species of highlymodified, often blind and scaleless forms that show numerousadaptations for cave life (see Caves). Theirisolated, easily disturbed habitats and bizarre appearance has made them vulnerable to both collecting and habitatdisruption, and several species are threatened and henceprotected nationally and internationally.
The gadiforms include some of the most important commercialfishes in the world such as the cods, haddocks, hakes, pollocks, and whitings (Fig. 14.25A). Gadiforms lacktrue spines but have experimented with fin rays. The longdorsal fin is relatively diversified compared with most primitivegroups; it is often divided into two or three parts, ananterior ray that is sometimes spinous (grenadiers) or elongateand even filamentous (morid cods, codlets, eel cods).True cods (Gadinae) have three dorsal fins and two anal fins.Pelvic fins are thoracic or jugular in position and are some

Fig. 14.24
Spawning behavior of the Pirate Perch, Aphrododerus sayanus, a fishwith a jugular-positioned anus. Pirate Perch spawn in dense root mats,the female and then the male pushing into narrow canals to depositeggs and sperm. Inset: a female in the process of laying eggs. FromFletcher et al. (2004), used with permission.
Many specieshave chin barbels (grenadiers, morid cods, eel cods, physidhakes, cods), a convergent trait in benthic or near-benthic (surpabenthic, benthopelagic) fishes. Gadiforms are northernmarine fishes with the solitary exception of the Burbot,Lota lota, which is a lake gadid of Holarctic (high latitude,northern hemisphere) distribution. The commerciallyimportant Atlantic Cod, Gadus morhua, is the largest speciesin the order; it once reached lengths of 1.8 m and weighedover 90 kg, although fish over 10 kg are now rare due to
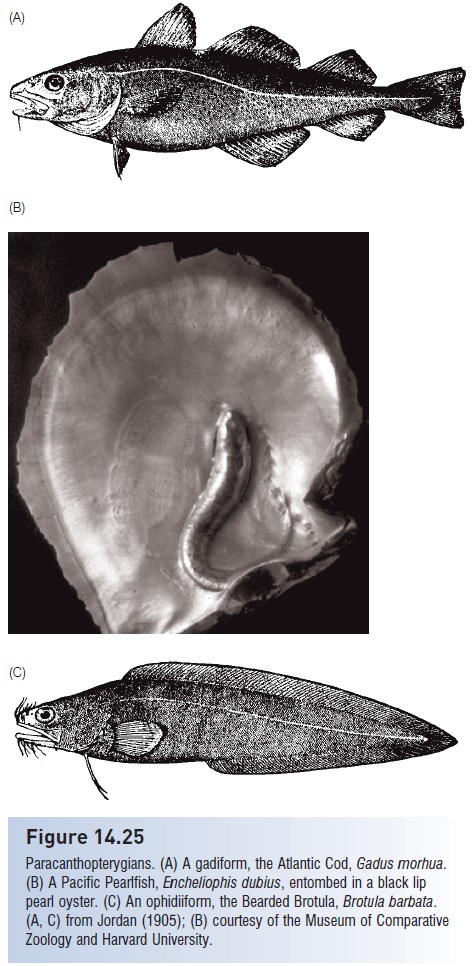
Figure 14.25
Paracanthopterygians. (A) A gadiform, the Atlantic Cod, Gadus morhua.(B) A Pacific Pearlfish, Encheliophis dubius, entombed in a black lippearl oyster. (C) An ophidiiform, the Bearded Brotula,Brotula barbata.(A, C) from Jordan (1905); (B) courtesy of the Museum of ComparativeZoology and Harvard University.
extensive overfishing. Previously “inexhaustible” cod fisherieshave crashed throughout much of the North Atlantic,affecting entire ocean ecosystems and the fishing economiesdependent on them (Mowat 1996; Kurlansky 1997). Thelargest food fishery in the world in recent times has beenfor North Pacific Walleye Pollock, Theragra chalcogramma,with a harvest in excess of 6 million tons in 1989.
In ophidiiforms the pectoral fins are high up on the bodyand have a vertical orientation. The pelvic fins, whenpresent, are located anteriorly under the head in what istermed the mental or jugular position. Pelvic fin loss in thisgroup probably relates to their eel-like bodies; many eellikefishes, regardless of taxonomic position, have reducedor absent pelvic fins and girdles (see Habitatuse and choice). Ophidiiforms inhabit what have to beviewed as marginal or at least exceptional habitats forfishes. Carapid pearlfishes are inquilines (tenants), livinginside the body cavities of starfishes, sea cucumbers, bivalves,and sea squirts; some may be parasitic, feeding on theinternal organs of their hosts. The common name originatedwith the discovery of the oyster’s revenge, wherean individual Pacific Pearlfish, Encheliophis dubius, becameentombed in a blacklip oyster (Pinctada mazatlantica) (Fig.14.24B). Pearlfishes are apparently unique among fishesin that they have two distinct larval stages, a “vexillifer”pelagic stage followed by a “tenuis” demersal stage duringwhich they search for a host. Ophidiid and bythitid cuskeelsand brotulas include blind species in freshwater cavesof the Caribbean basin and Galápagos Islands, and infaunalcoral reef species that hide deep within crevices (Fig.14.24C). The depth record for a fish is held by a neobythitinecusk-eel, Abyssobrotula galatheae, taken 8370 m downin the Puerto Rico Trench (see The deep sea).Bythitid brotulas and parabrotulid false brotulas are livebearers,a rare derivation among paracanthopterygians.
Batrachoidiforms are well-camoufl aged, benthic marinefishes with eyes placed high on the head, fl attened heads and large mouths, relatively elongate dorsal and anal fins,multiple lateral lines, and only three pairs of gills (ratherthan the usual five pairs). Their dorsal fins have two orthree stout spines. Midshipmen (Porichthys) have 600–800lateral photophores; photophores are unusual amongshallow water fishes. Fishes in this order are often quitevocal, producing a variety of sounds with their gas bladders.The muscles that vibrate the swim bladder of toadfishes arethe fastest contracting muscles known among vertebrates(Rome et al. 1999). Male midshipmen have been the focusof complaints by houseboat dwellers in San Francisco Bayduring the (midshipmen’s) breeding season, when the malesproduce a sustained, low-frequency “hum” (Ibara et al.1983). In the venomous toadfishes of the advanced subfamilyThalassophryninae, dorsal and opercular spines arepart of a complex system that injects a powerful venom.Toadfishes are unusual zoogeographically because they area shallow, warm water family that is most diverse in theAmericas, whereas most tropical marine families have theirgreatest diversity in the Indo-Australian region (Collette &Russo 1981; Marine zoogeographicregions). Three South American species are restricted tofresh water. In morphology, ecology, and venom production,toadfishes are convergent with the scorpaeniformstonefishes and perciform weeverfishes.
The most advanced order within the Paracanthopterygiiis the lophiiforms, a diverse and often bizarre-lookinggroup of marine fishes that are primitively benthic, shallowwater dwellers but that have evolved many highly modified,open water, deepsea forms. Many if not most of them usea modifiedfirst dorsal spine as a lure for catching smallerfish. The basal group is the lophiid goosefishes, knowncommercially as monkfish or poor man’s lobster. Goosefishes occur on both sides of the Atlantic and also in thePacific and Indian oceans. The western North AtlanticGoosefish, Lophius americanus, can exceed 1 m in lengthand 40 kg in mass, and has a huge mouth with long,recurved teeth that point back into the mouth. Goosefishesprey on other fishes and on diving seabirds. Antennariidfrogfishes also rest on the bottom and are wellcamouflaged, globose fishes that can walk across the bottomon their pectoral and pelvic fins (an old name for the lophiiforms,Pediculati, refers to the elbowlike bend in the pectoraland the footlike appearance of the pelvic fins). Theesca or lure of frogfishes can be quite ornate, mimicking asmall fish, shrimp, or worm (Fig. 14.26). When not wavedin front of potential prey, the esca sits in a protectivedepression between the second and third dorsal spines. Ifthe esca is bitten off, it apparently can regenerate back toits species-specific form (Pietsch & Grobecker 1987).

Figure 14.26
Lophiiformes. (A) A 6 cm long Bloody Frogfish, Antennariussanguineus, from the Galápagos Islands. Note the elaborate esca or lure at the end of the modified first dorsal spine or illicium. (B) Dorsalview of a Batfish, Dibranchus spinosa. Opercular openings are the teardropshaped holes at about midbody. (A) drawing by C. L. Starks, inHeller and Snodgrass (1903); (B) after Briggs (1974).
Geographically restricted fishes make up the family ofbrachionichthyid handfishes (Fig. 14.27). These small, colorful (red, orange, or pink with dark spots) benthicfishes occur only in southeastern Australia, with five of theeight known species restricted to Tasmania. Unlike the vastmajority of marine fishes, handfishes lack a pelagic larvalstage, fully formed juveniles emerging from eggs that areguarded by the female for 7–8 weeks. Because of their size,restricted range, attractive coloration, threats from introducedpredators, and low dispersal of both young andadults, most species are considered highly imperiled. Thespotted handfish, Brachionichthys hirsutus, has CriticallyEndangered status with IUCN.
The ogcocephalid batfishes are among the least fishlikefishes around. The fl attened, rounded head accounts for morethan half the length of the body; it tapers quickly behind theexpanded pectoral fins, giving the fish the appearance of arounded axe with a short handle. Batfishes alternate walkingon their pectorals with swimming via jet propulsion of waterexpelled from their round, backward-facing opercular openings.As modified as the batfishes are, they are rivaled instrange appearance by the 11 families of deepsea anglerfishes, suborder Ceratioidei (Fig. 14.28). The ceratioids arethe most speciose fishes of the vast bathypelagic region,comprising 160 species. Among their other derived traits,23 species in five families have very small males that fuse toand become parasitic on the larger females, the difference inlength between male and female being as much as 60-fold(Pietsch 2005; Pietsch & Orr 2007; Thedeep sea). Male parasitism appears to have evolved independentlyperhaps seven times in the suborder.

Figure 14.27
Brachionichthyid handfishes. A male and female Red Handfish(Brachionichthys politus) guarding their relatively large, yellow-brown
eggs. Handfishes get their name from their pectoral fins, which arearmlike appendages with an elbow and fingers. Handfishes are
restricted to southeastern Australian waters, chiefly Tasmania.
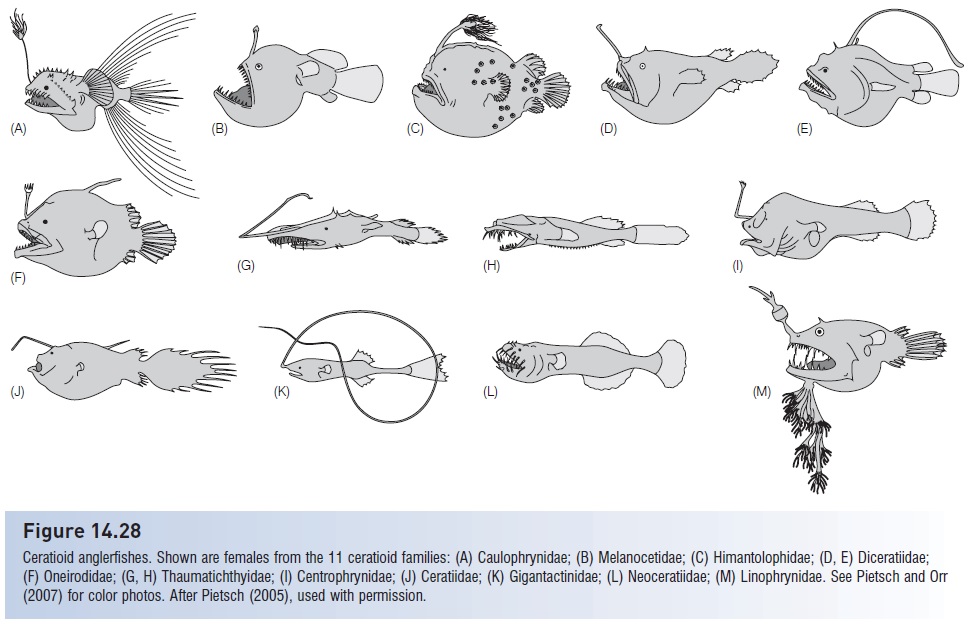
Figure 14.28
Ceratioid anglerfishes. Shown are females from the 11 ceratioid families: (A) Caulophrynidae; (B) Melanocetidae; (C) Himantolophidae; (D, E) Diceratiidae;(F) Oneirodidae; (G, H) Thaumatichthyidae; (I) Centrophrynidae; (J) Ceratiidae; (K) Gigantactinidae; (L) Neoceratiidae; (M) Linophrynidae. See Pietsch and Orr(2007) for color photos. After Pietsch (2005), used with permission.
Related Topics