Chapter: The Massage Connection ANATOMY AND PHYSIOLOGY : Endocrine System
The Pituitary Gland (Hypophysis) - The Endocrine Glands
THE PITUITARY GLAND (HYPOPHYSIS)
The pituitary gland is a small, oval gland that lies in a bony recess in the sphenoid bone at the base of the
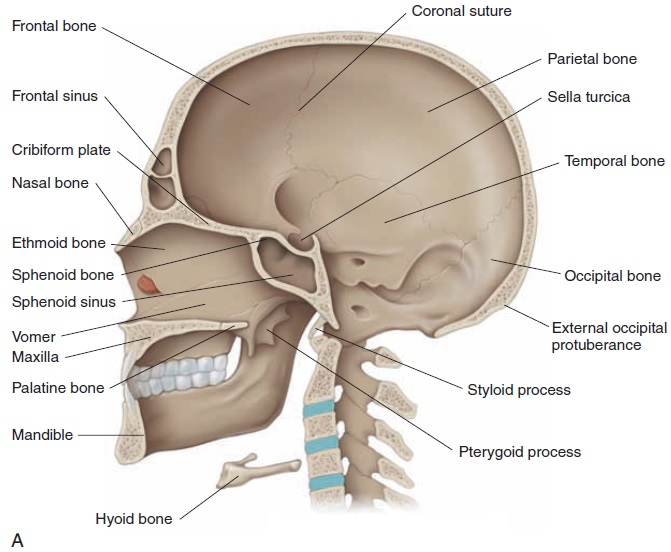
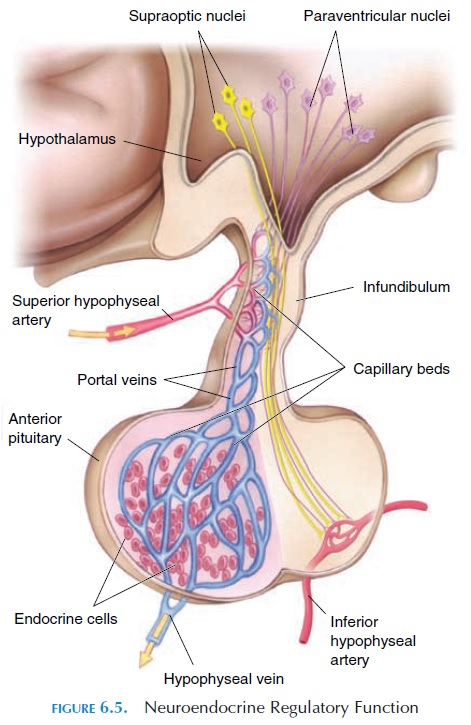
The gland, which is about the size of four peas (1–1.5 cm or 0.39–0.59 in), lies inferior to the hypothalamus con-nected by nerves and blood vessels (Figure 6.5). The pi-tuitary gland is divided into three lobes—anterior, in-termediate, and posterior. Because each lobe secretesdifferent hormones, the lobes can be considered as three separate endocrine organs that together secrete a total of 14 or more hormonally active substances.
Anterior Lobe (Adenohypophysis)
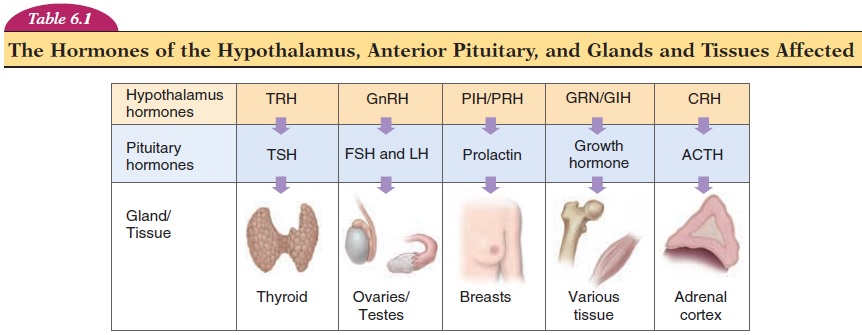
The anterior lobe is connected to the hypothalamus by special blood vessels known as the portal hypophyseal vessels. It secretes six established hormones—TSH, ACTH, LH, FSH, prolactin, and growth hormone.
The function of each of these hormones is self-explanatory. TSH controls thyroid gland secretions, ACTH controls the secretions of the adrenal cortex of the adrenal gland, LH (lutein, white) affects the corpus luteum formation in the ovaries, and FSH affects ovar-ian follicle development. Note that LH and FSH also have an important role in male reproductive function. Prolactin is one of the hormones needed for milk pro-duction, and growth hormone, together with many other hormones, affects growth. In addition, the ante-rior pituitary also secretes ß-lipotropin, which con-tains the amino acid sequences of endorphins and enkephalins.
Cells that secrete the hormones manufacture and store the hormones in vesicles. When stimulated, the cells secrete the stored hormones into the blood where they circulate throughout the body to reach the target organs.
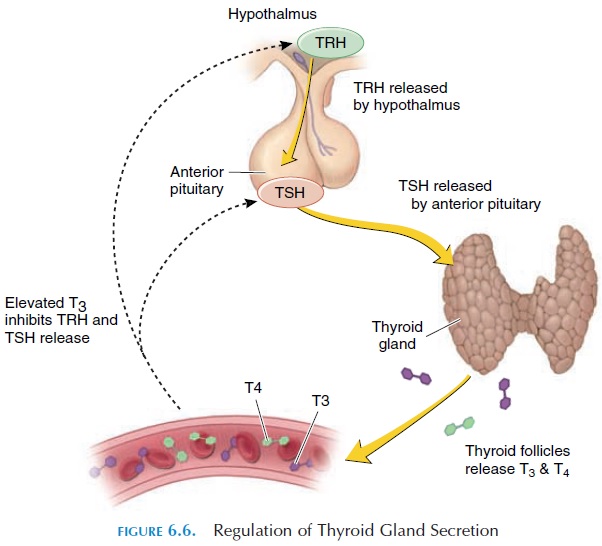
Thyroid Stimulating Hormone (TSH) (Thyrotropin)
Alterations of the circulating levels of TSH regulate the secretion of hormones from the thyroid gland. TSH levels are, in turn, controlled by TRH of the hy-pothalamus (Table 6.1). By negative feedback mecha-nism, increasing levels of thyroid hormone in the cir-culation reduce TRH and TSH secretion (see Figure 6.6). Within a few hours of secretion, TSH is rapidly removed from circulation by the kidney and liver. Prolonged thyroid stimulation by TSH can result in an enlargement of the thyroid called agoiter (see Ab-normalities of Thyroid Secretion). Abnormalities of TSH secretion by the pituitary can, therefore, present as hyposecretion or hypersecretion of the thyroid.
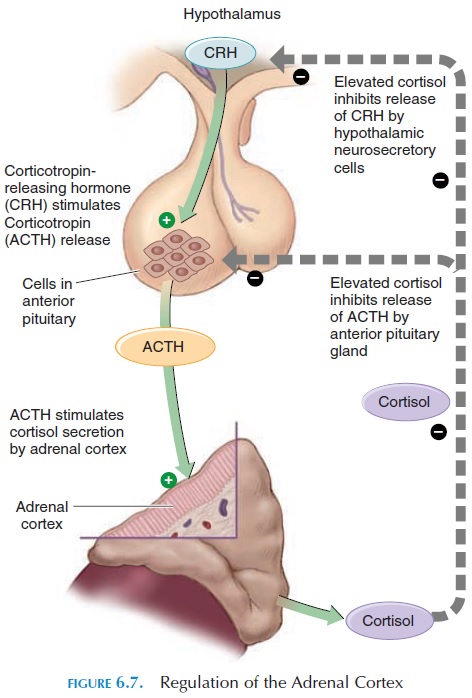
Adrenocorticotropic Hormone (ACTH)
The cells in the adrenal cortex that produce glucocor-ticoids have receptors for ACTH. Binding of ACTH to these receptors results in steroid synthesis. The level of steroids regulates ACTH secretion (see Figure 6.7) by negative feedback mechanism.
Luteinizing Hormone (LH) and Follicle-StimulatingHormone (FSH)
LH stimulates ovulation and formation of corpus lu-teum from female ovarian follicles and stimulates male testosterone secretion. FSH stimulates the growth of female ovarian follicles and the formation of sperm (spermatogenesis) in males. Absence of pi-tuitary hormones can result in atrophy of the testis and ovaries.
Prolactin
Prolactin initiates and maintains breast milk secre-tion after the breasts have been primed by the hor-mones estrogen, progesterone, glucocorticoids, hu-man growth hormone, thyroxine, and insulin. It also inhibits the effects of FSH and LH on the ovaries and is probably responsible for the slow onset of men-strual cycles in lactating women. Although prolactin is required for milk production, the ejection of milk is initiated by the hormone oxytocin. The role of pro-lactin in males is not fully understood.
Abnormal prolactin levels can result in milk pro-duction, even in the absence of pregnancy (galactor-rhea) and absence of menstruation (amenorrhea).
Hypersecretion of prolactin has been associated with impotence in men.
Growth Hormone
Growth hormone is a protein hormone that, as the name suggests, affects growth. It increases the for-mation of cartilage in bone and increases height in young children in whom the epiphysis is not fused. Other than affecting growth, this hormone increases glucose levels in blood and collagen formation.
The effects of growth hormone on growth, carti-lage, and protein anabolism depend on its interaction with other growth factors (proteins) called so-matomedins produced by the liver, skeletal muscle,cartilage, bone, and other tissue. A variety of growth factors have been identified, such as insulinlikegrowth factor, nerve growth factor, epidermal growth factor, ovarian growth factor, and fibroblast growth factor (depending on the tissue and organ they af-fect). Growth factors cause cells to grow and multiply by increasing the uptake of amino acids and decreas-ing the breakdown of proteins. Growth factors in-crease the breakdown of fat from adipose tissue; the fatty acids released are used for energy production. They decrease the uptake of glucose by cells, sparing glucose for utilization by nervous tissue. They also stimulate the release of glucose from the liver into the blood by the breakdown of liver glycogen.
Other than growth hormone, individual growth is affected by thyroid hormones, sex hormones, steroids, and insulin. Genetic factors and adequate nutrition also play an important role in growth.
Growth hormone secretion is primarily controlled by growth hormone-inhibiting hormone (GHIH) and growth hormone-releasing hormone (GHRH) of the hypothalamus, which are then regulated by glucose levels in the blood. Other factors also affect growth hormone secretion, such as levels of fatty and amino acids; increased activity of the sympathetic nervous system; and hormones, such as glucagon, insulin, es-trogen, and cortisol.
Intermediate Lobe
The intermediate lobe is rudimentary in humans and appears to secrete hormones in insignificant levels. In humans, some of the cells migrate to the anterior pi-tuitary and secrete the hormones melanocyte-stimu-lating hormone (MSH) and endorphins. The physi-ologic function of MSH in humans is not known; however, in lower animals, such as fish, reptiles and amphibians, it results in darkening of the skin. The darkening of the skin when these animals are placed in a dark background is another example of a neu-roendocrine reflex, in which nerves from the eye alter endocrine secretions in accordance to the external environment.
In mammals, treatment with MSH increases syn-thesis of melanin and darkens the skin of humans in 24 hours. Because ACTH has some MSH activity, ab-normally high levels of ACTH can result in darkening of skin creases and other areas of the body.
Endorphins are small peptides found, in addition to the pituitary gland, in many parts of the body, in-cluding the nervous system. With other peptides— enkephalins and dynorphins—endorphins were iden-tified when scientists discovered that brain cells had receptors for morphine, a synthetic painkiller. These peptides are thought to be the natural painkillers of the human body and are, therefore, known as opioid peptides or endogenous opioids. Two hundred times more potent than morphine as analgesics, these pep-tides have been linked to memory and learning, plea-sure sensations, body temperature control, regula-tion of hormones that affect onset of puberty, sexual drive and reproduction, and disorders such as de-pression and schizophrenia.
Posterior Lobe (Neurohypophysis)
The posterior pituitary does not have its own secre-tory cells. Instead, it has the endings of axons whose cell bodies lie in the hypothalamus (Figure 6.4). These axons secrete oxytocin and vasopressin (an-tidiuretic hormone [ADH]) into the capillary net-work located in this region. The hormones secreted by the posterior pituitary are typically neural hor-mones—hormones secreted into the circulation bynerve endings.
Oxytocin
The primary target organs of oxytocin are the uterus and breasts (see Figure 6.8). In mammals, nipple stimulation results in the generation of impulses in sensory nerves located in and around the nipples. These impulses communicate with the nerve cells in the hypothalamus that manufacture oxytocin. Oxy-tocin, released by the axons of these nerve cells into the capillaries in the posterior pituitary, is trans-ported to the breasts where it stimulates contraction of smooth muscle cells. The smooth muscle cells (myoepithelial cells) located around the ducts of the breast contract and eject the milk that has already been produced. This is known as the milk ejectionreflex, or milk let-down reflex, and is an example ofa neuroendocrine reflex.
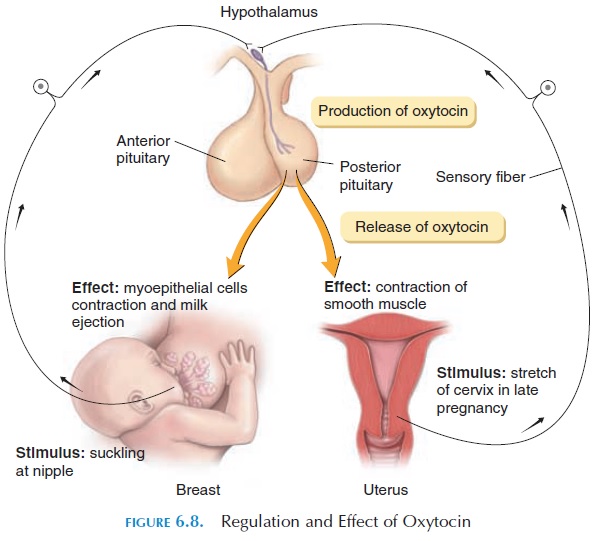
In addition to its action on the breasts, oxytocin causes contraction of the smooth muscle of the uterus. In late pregnancy, the uterus becomes more sensitive to oxytocin as a result of an increase of oxytocin re-ceptors on the smooth muscle cells (up-regulation). Stretching of the cervix by the descending fetus results in an increase in impulses in the sensory nerves lo-cated in this region.These sensory nerves communi-cate with the hypothalamus and increase production of oxytocin. During labor, as the fetus further dilates the cervix, further increase in oxytocin levels result, producing strong contractions of the uterus. This con-tinues until the fetus is expelled and is one of the few situations in the body where positive feedback occurs. Oxytocin, however, is not the only hormone or mecha-nism involved in labor.
Oxytocin may also play a part in the transport of sperm up the reproductive tract at intercourse and the feeling of sexual pleasure during and after inter-course. In men, oxytocin stimulates the contraction of the vas deferens and the prostate glands before ejaculation occurs. In animal experiments, this hor-mone has been linked to parental bonding with off-spring.
Vasopressin or Antidiuretic Hormone (ADH)
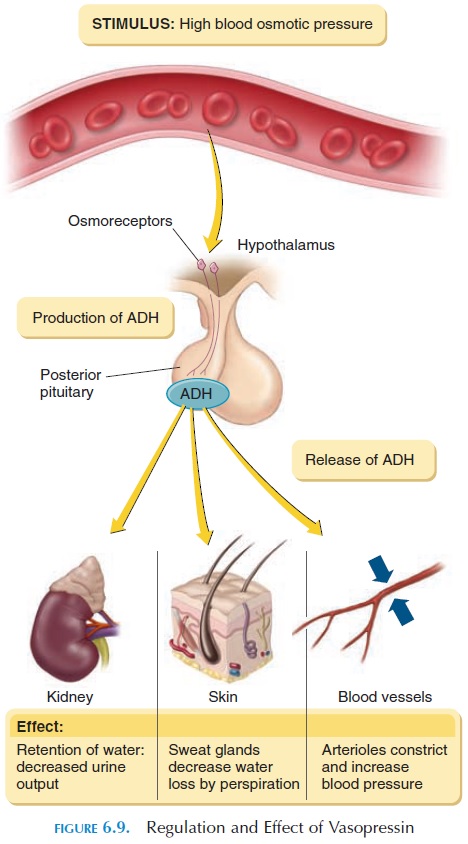
A major function of vasopressin is to conserve water by acting on the kidney; hence, the name antidiuretic hormone. Vasopressin acts on the collecting ducts of the kidney , making them more perme-able to water. The water moves into the interstitial re-gion from the ducts, reducing urine volume and in-creasing urine concentration. In the absence of vasopressin, more dilute urine is formed. ADH also decreases the loss of water through sweat.
Vasopressin in large doses causes contraction of the smooth muscle of the arterial walls and a rise in blood pressure. This regulatory mechanism is partic-ularly useful when blood pressure drops after hemor-rhage. Circulating vasopressin is rapidly removed by the kidneys and the liver; the half-life of vasopressin is about 18 minutes.
Vasopressin secretion is triggered by stimulation of osmoreceptors located in the hypothalamus. These receptors respond to changes in osmotic pres-sure in the blood. When the osmotic pressure in plasma is increased, the extracellular fluid volume is decreased, or when a person stands up from lying down, vasopressin levels increase to reverse the situ-ation (see Figure 6.9).
Related Topics