Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: Teleosts at last I: bonytongues through anglerfishes
Subdivision Euteleostei - teleostean fishes
Subdivision Euteleostei
All the teleosts above the level of the ostariophysans aregenerally placed together in the subdivision Euteleostei, the“true” teleosts. This designation underscores the conclusionthat teleostean phylogeny involved four major radiations,the first three producing relatively primitive andseparate groups (osteoglossomorphs, elopomorphs, otocephalans),and the fourth containing a vast (346 families,2935 genera, 17,419 species) assemblage of more advanced,euteleostean fishes.
The monophyly of the euteleosts as well as the organizationof the subdivision remain an area of debate becauseunique shared derived characters common to all or mostmembers are lacking (or are confused by exceptions), andevidence from different approaches (e.g., anatomical,developmental, and molecular) supports differing conclusions.Johnson and Patterson (1996) proposed a scheme,followed here, that emphasizes three shared traits:
1 Similarities in the pattern of embryonic developmentof the supraneural bones among euteleosts, which arethe small T-shaped or rodlike bony or cartilaginouselements that lie within the musculature between thecranium and the dorsal fin.
2 The presence of an outgrowth on the stegural bone, astructure associated with the neural arches of thevertebral centra of the tail base.
3 The presence of caudal median cartilages, cartilagesthat lie between the hypural bones of the caudal base.
Johnson and Patterson’s analysis eliminates reliance oneven more problematic structures for defining the euteleosts.Older summaries emphasized the shared existence ofstructures that had probably evolved convergently among taxa or that were shared with groups clearly primitive toeuteleosts, or that were missing from important clades. Onesuch trait was the presence of epidermal breeding turberclesand dermal contact organs, small bumps on the fins andbodies that develop during the breeding season in 25 families(Collette 1977; Sexual selection,dimorphism, and mate choice). Another, highly enigmatictrait was the presence of anadipose fin. This small, (usually)rayless fin sits posterior to the first dorsal, often just anteriorto the caudal peduncle. It can be a small vertical flapor bump or it can be a long, substantial structure confl uentwith the caudal fin, as in many catfishes. It occurs in characiforms,catfishes, smelts, the deepsea stomiiforms, aulopiforms,and scopelomorphs, the salmoniforms (salmons,trouts, whitefishes), and trout-perches. It is generally lacking

Figure 14.14
Gymnotiform electric knifefishes. An accurate rendering ofelectric eels, Electrophorus electricus, in an otherwisedramatized setting. Anonymous source.from higher fishes, particularly those with a spiny firstdorsal. Its functions are a mystery.
Subdivision Euteostei,Superorder Protacanthopterygii
Protacanthopterygians as a group have undergone repeatedrevision, taxa being removed and added to the superorderas new information is obtained and old data are reinterpreted.The name is used here because it provides an organizationalcategory for a series of apparently related fishes(Fig. 14.15). Nelson (2006) retained Osmeriformes as aseparate order within this superorder; we have chosen tofollow Johnson and Patterson (1996) and considerosmeroids a suborder within the salmoniforms. We havesimilarly chosen to separate the esociform pikes and mudminnowsfrom the Protacanthopterygii, aligning them withmore advanced teleostean groups (see below, Neognathi).

Figure 14.15
Protacanthopterygians. (A) An osmerid, the Capelin,Mallotus villosus (sexually mature female). (B) TheLongfin Svetovidov’s Char, Salvethymus svetovidovi,of Lake El’gygytgyn, Siberia. This long-lived, smallchar (30 years, 30 cm) is threatened by pollutionand fishing. (C) A large Chinook Salmon,Onchorhynchus tshawytscha, from British Columbia.(A) from Bigelow (1963), used with permission;(B) drawing by Paul Vecsei, used with permission;(C) photo courtesy of R. Carlson.
Argentiniforms are primarily deepsea inhabitants. The barreleyes are bizarre looking deepseafishes with elongate tubular eyes that point upwards. Theplatytroctids are also deepsea fishes that exude a blue-greenluminous fluid from a papilla located under the shouldergirdle, perhaps analogous to squid ink; many species alsopossess photophores (light organs), a convergent traitamong deepsea species.
Within the Salmoniformes are two suborders, theSalmonoidei and the Osmeroidei. Salmonoids are an important group commercially, ecologically, and esthetically, anda fascinating group evolutionarily. They are the focusworldwide of fisheries classes and of popular and technicalbooks. Different taxonomic treatments recognize as manyas three different families (coregonid whitefishes, thymallidgraylings, salmonid salmons and relatives), but we followNelson (2006) and treat them as subfamilies of a largerSalmonidae. Their general anatomical similarities includean adipose fin, no spiny fin rays, a triangular fl ap at thebase of the pelvic fin (pelvic axillary process), gill membranesfree from the ventral side of the head, maxillaincluded in the gape, a physostomous gas bladder, andvertical barring (parr marks) on the sides of most young.Internally the last three vertebrae angle up toward the tail,and a myodome, or area of the skull where the extrinsiceye muscles insert, is present.
The coregonine whitefishes and ciscoes consist ofapproximately 32 species of relatively large-scaled salmonids that lack teeth on the maxillary bone.
They arezooplanktivorous fishes in high-latitude lakes of NorthAmerica and Eurasia that show a great deal of withinspeciesvariation and specialization, forming species complexesthat differ from lake to lake. Kottelat (1997)documented at least five apparent extinctions and severalmore extirpations of unique coregonines in Europe. Duringthe mid-20th century, two coregonines native to theLaurentian Great Lakes, the Deep Water Cisco, Coregonusjohannae, and the Blackfin Cisco, C. nigripinnis, disappearedas the result of overexploitation, pollution, siltation,competition with nonindigenous species, and perhaps predationby introduced Sea Lampreys. Whitefishes exemplifythe vulnerability of taxa that develop localized specializations(the Wild Salmon Center and Ecotrust maintains a website at www.stateofthesalmon.org that includes accountsof imperiled salmonids based on information from theInternational Union for the Conservation of Nature(IUCN)’s Salmon Specialist Group). The thymalline graylings,Thymallus, are a smaller group of about five speciesof northern hemisphere riverine salmonids that are easilyidentified by an elongate, flowing dorsal fin.
The subfamily Salmoninae contains seven Eurasian andNorth American genera (Brachymystax, Acantholingua,
Salmothymus, Hucho, Salvelinus, Salmo, Oncorhynchus)that differ from other salmonids by having small dorsal fins,small scales, and teeth on the maxillary bone (Fig. 14.15B,C). Most species of economic importance are in the latterthree genera, although the Siberian Taimen, Hucho taimen,is the world’s largest salmonid at 2 m and 70 kg (a commerciallycaught individual weighed 114 kg). North AmericanSalmoninae are currently divided into three genera andapproximately 20 species, the names and relationships ofwhich have been the subject of considerable debate. The chars (or charrs) include the Lake,Brook, and Bull trouts, Arctic Char, and Dolly Varden, allin the genus Salvelinus. The northernmost-living freshwaterfish in the world is the Arctic Char, Salvelinus alpinus, of


Figure 14.16
Phylogeny of the salmonids. A cladogram of most living salmonids based on life history traits shows the evolution of the various species. Thesame cladogram is constructed if anatomical and biochemical traits are used. The first four lineages represent separate genera (Thymallus,Brachymystax, Hucho,Salvelinus), Atlantic Salmon and Brown Trout are in the genus Salmo, and the remaining eight species (those above trait4) are all in the genusOncorhynchus. The more primitive coregonine whitefishes would come off to the far left of the cladogram and are notshown. The following life history and reproductive characteristics are the shared derived characters that were used to construct the cladogram.The listed characters correspond to the numbered branch points in the cladogram. Groups to the right and above the number possess the trait,those to the left do not. From Smith and Stearly (1989), used with permission.1, egg diameter greater than 4.5 mm; females dig redds (nests); large males have hooked jaws (kype) during breeding season; 2, fallspawners; 3, commonly undergo long oceanic migrations; 4, most spawners undergo irreversible hormonal changes; 5, spring spawners; 6,anadromous forms die after spawning; 7, non-migratory individuals tend not to reproduce; 8, most smolt in first year, some go to sea as evenyounger fry; 9, juveniles are strong schoolers, parr are slender; 10, the freshwater phase is reduced; young migrate soon after emerging fromgravel.
▲
The remaining salmonines are the Atlantic basin salmonand trout (Salmo salar, the Atlantic Salmon; S. trutta, the European Brown Trout, with numerous subspecies andraces; and three other European species), and the 11 species of Pacific basin trouts and salmons in the genus Oncorhynchus(Behnke 2002) (Fig. 14.17). Two Oncorhynchusspecies, O. masou and O. rhodurus, are endemic to Japan.Pacific trouts and salmons include narrowly distributed,landlocked forms such as Golden and Gila trouts, andspecies that are spectacularly anadromous, such as theChinook, Chum, Coho, Pink, and Sockeye salmons (O.tshawytscha, O. keta, O. kisutch, O. gorbuscha, and O.nerka, respectively). (Latinized names of Pacific salmon defy the usually predictable logic of binomial nomenclature.The mystery is partially solved when one learns thatthese spellings represent Russian names transliterated intoEnglish and later converted into Latin over a 60-year periodby a series of German and British ichthyologists (Moyle2002).) Some Pacific salmon undergo oceanic migrations ofthousands of kilometers before returning to their birth riverto spawn and die (Representative life historiesof migratory fishes). The actual number of species,subspecies, and distinct races of Pacific salmons is a matterof considerable and important debate because of the wholesaledestruction of stocks in various rivers of the PacificNorthwest region of the USA (Lichatowich 1999; Williams2006). Many of these stocks are reproductively isolated andgenetically distinct and therefore are viewed as unique evolutionarytaxa, termed evolutionary significant units (ESUs)for conservation purposes.

Figure 14.17
A Golden Trout, Oncorhynchus aguabonita, the state fish of California.Greatly depleted by introduced species and habitat destruction, GoldenTrout are now the focus of restoration programs that include eradicationof the European Brown Trout that were actively stocked in pastdecades. Photo by T. Kelsey, courtesy of E. P. Pister.
In the osmeroids is the family Osmeridae, which includesabout 30 species of marine, freshwater, and diadromous (migrating between fresh water and the sea) species thatinhabit shallow waters. Osmerids are generally small,silvery, elongate fishes that swim in the water column. Theyhave a single, soft-rayed dorsal fin and sometimes an adiposefin. The maxilla is included in the gape, and most jaw bonespossess teeth. Although their pelvic fins are abdominal, asis the case for most of the preceding teleostean groups,some osmerids have their pectorals located higher on thebody than is common in the more primitive groups.Osmerids include commercially important species such asCapelins (Mallotus), Eulachons (Thaleichthys), Asian Ayu(Plecoglossus), and Rainbow Smelt (Osmerus), many ofwhich are superfi cially similar to the more advanced silversides(Atherinidae).
The other osmeroids are two families that dominate coldfreshwater environments of the southern hemisphere. Theseare the retropinnids and galaxiids. Retropinnids are small(10–35 cm), marine and anadromous (spawn in fresh water, grow in the sea) fishes of New Zealand and Australia knownas southern smelts and southern graylings; they sometimesestablish landlocked populations in lakes. The galaxiidsoccur in Australia and New Zealand but also in southernSouth America and Africa. Galaxiids constitute importantcommercial whitebait fisheries in New Zealand. Galaxiidshave complex life cycles, exhibiting all major types ofdiadromy, including anadromy, catadromy, and amphidromy(see Diadromy). Some are also semelparous,spawning only one time before dying (see Life histories and reproductive ecology). Retropinnidsand galaxiids as a group have suffered numerous extirpationsand extinctions as a result of habitat destruction andthe stocking of non-native trouts (see Speciesintroductions). One retropinnid,Prototroctes oxyrhinchus,is known to be extinct.
A truly unique galaxiid is the salamanderfishLepidogalaxiassalamandroides (Fig. 14.18). This benthic living, elongatefish inhabits seasonally dry ponds of southwesternAustralia, where it buries in the mud and lives in a torpidstate after ponds dry up, re-emerging with the next rains(see Deserts and other seasonally arid habitats).Salamanderfish lack eye muscles but instead have a fl exibleneck joint that allows them to bend their neck at a right angleto the side, a very unusual ability in fishes. Neck bending ismade possible by large gaps between the back of the skulland the first cervical vertebrae and between the first andsecond cervical vertebrae. Salamanderfish have also apparentlyreinvented the physostomous gas bladder, in that theylack a normal gas bladder but instead have a gas-containingstructure made up of simpler mesentery-like tissue and collagenfi bers (Berra & Allen 1989; Berra et al. 1989).
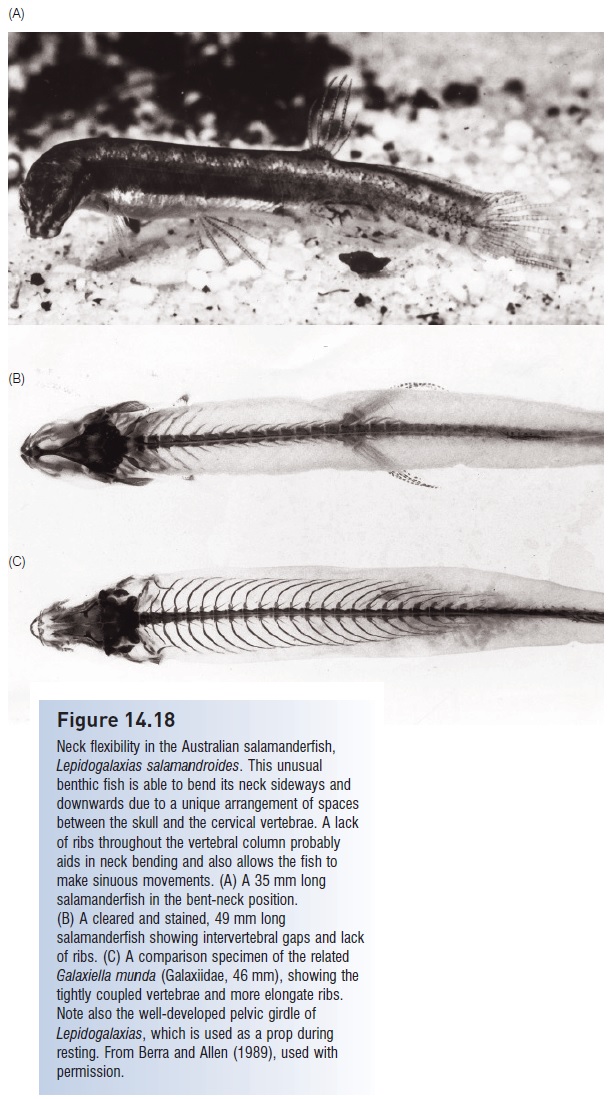
Figure 14.18
Neck flexibility in the Australian salamanderfish,Lepidogalaxias salamandroides. This unusualbenthic fish is able to bend its neck sideways anddownwards due to a unique arrangement of spacesbetween the skull and the cervical vertebrae. A lackof ribs throughout the vertebral column probablyaids in neck bending and also allows the fish tomake sinuous movements. (A) A 35 mm long salamanderfish in the bent-neck position.(B) A cleared and stained, 49 mm longsalamanderfish showing intervertebral gaps and lack of ribs. (C) A comparison specimen of the relatedGalaxiella munda (Galaxiidae, 46 mm), showing thetightly coupled vertebrae and more elongate ribs.Note also the well-developed pelvic girdle ofLepidogalaxias, which is used as a prop duringresting. From Berra and Allen (1989), used withpermission.
Related Topics