Chapter: Plant Anatomy:An Applied Approach: The stem
Transport phloem within the axial system - Stems
Transport phloem within the axial system
The plant vascular system plays a pivotal role in the delivery of nutrients to distantly located organs. Recent discoveries have provided new insight into a novel role for plasmodesmata and the phloem in terms of the transport and delivery of information macromolecules including proteins and ribonucleoprotein complexes. Ruiz-Medrano et al. (2001) suggest that the
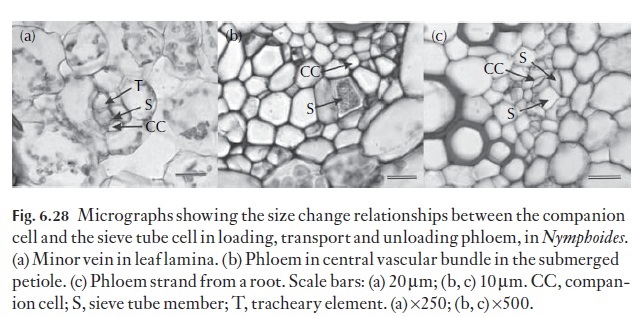
Within stems, a great deal of assimilate is transported through the axial system, to local or distant sinks. Figure 6.28 illustrates the difference in size between loading transport and unloading phloem in Nymphoides. Within the trans-port phloem, sieve tubes are between 10 and 25 µm in diameter, and their associated companion cells between 15 and 38 µm in diameter. Clearly, sieve tubes in transport phloem are larger than corresponding sieve tubes in loading phloem in this species.
The course of vascular systems in monocotyledonous stems has been studied for many years and is under active investigation at present. Modern microscopy techniques, including fluorescence and confocal microscopy, stack frame imaging of whole and sectioned material have enabled research-ers to understand for the first time the true complexity of many stems, in-cluding palms and Pandanaceae. With newer and more powerful techniques becoming available, a new area of comparative anatomy is emerging in the study of whole vascular systems. The results of this study might well show basic types which underlie the major phylogenetic divisions in the plant kingdom.
Within the vascular bundles, the phloem and xylem of primary systems show only axial systems of cells. The rays are a feature of secondary development.
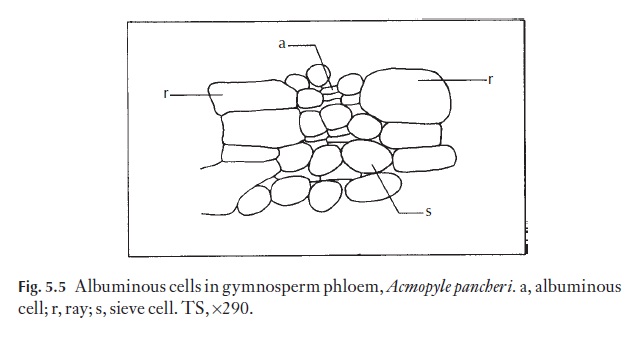
Phloem in gymnosperms has well-developed sieve cells, with sieve areas and their associated albuminous cells. In angiosperms the albuminous cells are replaced by companion cells (Fig. 5.5). It is thought by those who study phloem that an evolutionary sequence can be observed, from systems in which companion cells are poorly defined and the sieve tube elements communicate by rather scattered sieve areas on oblique walls to the most advanced in which sieve plates are very well defined and constitute the transverse end wall between elements in a sieve tube, and in which the com-panion cells are very well developed. Since the advanced sieve tube member
has no nucleus, the organization of the element is carried out by the nucleate companion cell adjacent to it. Damage to the companion cell in this system may bring about failure of the element which it directs. Phloem is not the easiest of tissues to study with the light microscope, and consequently it is only in recent years that the beautiful comparative studies such as those carried out in the laboratories of Katherine Esau and later by Ray Evert have contributed greatly to our understanding of the anatomy and struc-ture of phloem.
![]()
![]() Sieve plates can be seen easily in a number of plants, especially those that have large-diameter sieve tubes; a good example is the Cucurbitaceae.
Sieve plates can be seen easily in a number of plants, especially those that have large-diameter sieve tubes; a good example is the Cucurbitaceae.
As in the leaf, the first formed phloem in stems is termed protophloem. It is often functional for a short period only, as it usually differentiates in re-gions of rapid cell expansion and elongation and often becomes compressed, but not before the later-developing metaphloem matures.
Related Topics