Chapter: Plant Anatomy:An Applied Approach: Morphology and tissue systems: the integrated plant body
The transport systems - Morphology and tissue systems
The transport systems
It is not possible to present a simple, comprehensive model to demonstrate the wide range of arrangements of vascular systems that occur in vascular plants, or in either dicotyledons or monocotyledons for that matter. Dicot-yledons that are composed of wholly primary tissues tend to be a little more stereotyped than monocotyledons, but even then there is a very wide range of arrangements.
The essential elements of both systems are the xylem, concerned with transport of water and dissolved salts, and the phloem, which translocates synthesized but soluble materials around the plant to places of active growth or regions of use or storage. Xylem strands and phloem strands are normally associated and together form the vascular bundles, and are often enclosed in a sheath of fibres, and in addition, in some instances, an outer sheath of parenchyma cells (the bundle sheaths). Vascular bundles make up the ‘plumbing system’ of primary tissues, and organs without secondary growth in thickness.
In the apex (tip) of the shoot and root, where vascular tissue is not yet de-veloped, soluble materials and water move from cell to cell through special-ized very fine strands of protoplasm (called plasmodesmata) in these relatively unspecialized zones. Not far back from these growing points, however, more formal conducting systems are needed to cope with the flow of assimilate and water. Procambial strands, strands of elongated, thin-walled cells which are the precursors of the vascular bundles, are seen first and then, further from the tips, differentiation of protophloem (first formed primary phloem) alone followed by protoxylem (first formed primary xylem) and then by the metaphloem and metaxylem (next formed phloem and xylem cells respectively). The protoxylem and metaxylem, protoph-loem and metaphloem together constitute the primary vascular tissues. In most dicotyledons, the newly formed strands join the previously formed vascular bundles in the stem through a leaf or branch gap, which is com-posed of parenchyma cells, and ‘breaches’ the harder tissues associated with the plumbing of the stem.
In most dicotyledons the leaf lamina (blade) has a midrib to which are connected the lateral veins. The latter form a network composed of major and minor systems. The midrib is directly connected to the petiole trace, the vascular system of the petiole or leaf stalk. This enters the stem and joins into the main stem system through a leaf trace gap as described above. In the primary stem, all vascular bundles are separate from one anotherexcept at the nodes – those parts of the stem where one or more leaves are attached. Vascular bundles in the stem may remain separate in many climb-ers, e.g. Cucurbita, Ecballium, but in most dicotyledons the bundles become joined into a cylinder by growth of secondary xylem and phloem from vas-cular cambium (a lateral meristem composed of thin-walled cells from which the secondary vascular tissues develop); it is made up of the fascicular cambium forming within the vascular bundle and the interfascicular cam-bium between vascular bundles.
![]()
![]() A complex rearrangement of tissues takes place in the primary plant where the systems of the stem and root meet (hypocotyl). In the stem vascular bundles, the phloem is normally to the outer side of the xylem in the majority of plants. In the root, as seen in cross-section, the xylem is central, and may have several lobes or poles, with the phloem situated between these. After secondary growth has taken place, the hypocotyl becomes surrounded by secondary xylem and phloem, and the shoot and root anatomy become more similar.
A complex rearrangement of tissues takes place in the primary plant where the systems of the stem and root meet (hypocotyl). In the stem vascular bundles, the phloem is normally to the outer side of the xylem in the majority of plants. In the root, as seen in cross-section, the xylem is central, and may have several lobes or poles, with the phloem situated between these. After secondary growth has taken place, the hypocotyl becomes surrounded by secondary xylem and phloem, and the shoot and root anatomy become more similar.
Transfer cells are specialized parenchymatous cells found in various parts of the plant, but in particular, in regions where there is a physiological demand for transport, but where more normal phloem or xylem cells are not in evidence. A good example is the junction between cotyledons (first seedling leaves) and the shoot axis in seedlings. Transfer cells may also be present near the extremities of veins, or near to adventitious buds (buds developing in an unusual position, e.g. on a stem in addition to or replac-ing those in leaf axils, or buds on root or leaf cuttings).
Thin sections of the walls of transfer cells show them to have numerous small projections directed towards the cell lumen (the part of the cell to the inner side of and enclosed by the cell walls). These greatly increase the plasmalemma–cell wall surface interface. a site of metabolic activity con-cerned with the rapid, energy-mediated movement of materials between adjacent cells. The projections are so fine that conventional sections with a rotary microtome are too thick for them to be seen.
Monocotyledons are quite different from dicotyledons in their vascula-ture. Leaf and stem are commonly much less readily separable as distinct organs. There is no secondary growth by a true vascular cambium, so a cyl-inder of vascular tissue does not form. When secondary growth occurs, as in Dracaena and Cordyline, it is by means of specialized tissue, situated near to the stem surface, which forms complete, individual vascular strands and additional ground tissue.
Vascular bundles are usually arranged in the stem with the xylem pole facing towards the stem centre (but this is not invariably so). The arrange-ment of leaf vascular bundles is very variable. Grasses and some Juncus spe-cies, for example, often have one row as in Fig. 1.3. Some of the other types of arrangement are discussed.
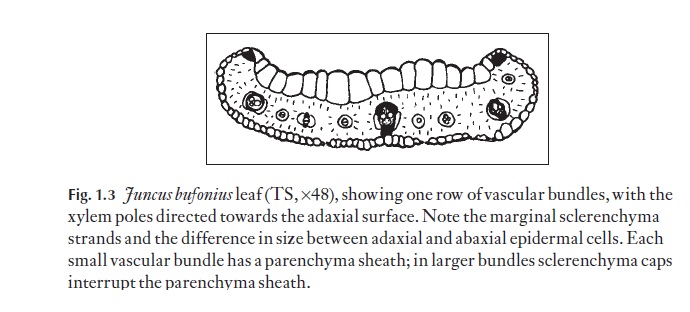
Because there is no vascular cylinder in monocotyledons, where leaf traces (bundles) enter the stem they do not form gaps. They may join at nodes, where all the bundles at that particular level of the stem form a type of plexus, as in aloes. Sometimes, in stems with nodes, the leaf traces may continue downwards from their points of entry into the stem for a complete internode before joining the nodal plexus below (e.g. Restio, Leptocarpus, Restionaceae). In other plants without nodes (e.g. palms), the leaf traces fol-low a simple path curving inwards towards the stem centre, and then gradu-ally ‘move’ towards the outer region of the stem lower down. These leaf traces join onto the main bundles by small, inconspicuous bridging bun-dles. This system is beautiful in its simplicity, but very difficult to analyse because there are so many (several hundred) vascular bundles even in the narrow portion of a stem of a small palm like Rhapis. As one follows the course of bundles in a palm, they are seen to spiral down the stem.
The primary root does not develop in a majority of monocotyledons. Its function is usually taken over by numerous adventitious roots that arise at an early stage, usually at the nodes, and join the stem vascular system in what frequently appears as a jumble of vascular tissue with very short ele-ments both in the phloem and xylem.
Related Topics