Chapter: Basic Concept of Biotechnology : Biomolecules
Structure of lipids
Structure of lipids
The structure of all three types of lipids is briefly discussed below.
Fatty Acids
Fatty acids consist of a long carbon chain (also called an acyl chain) with carboxylic acid at one end. The vast majority of fatty acids are unbranched linear molecules. The carboxylic acid is ionized at physiological pH (the carboxyl group is deprotonated and therefore negatively charged). Fatty acids in most biological systems are synthesized by serial addition of two carbon units. As a result, fatty acids usually contain an even number of carbons, especially in animals, which synthesize even-chain fatty acids almost exclusively. Biological fatty acids usually contain 14 to 20 carbons, although small amounts of 22 and 24 carbon compounds are found in some tissues. The fatty acid acyl chain “prefers” to be extended, because this results in the least steric hindrance; however, the chain is very flexible, and will adopt a large variety of conformations. The reason for this flexibility is that each carbon-carbon bond can (more or less) freely rotate, and all fatty acids have many carbon-carbon bonds.
Saturated fat and unsaturated fat
Many fatty acids contain double bonds. Fatty acids that do not contain carbon-carbon double bonds are considered to be saturated. When producing unsaturated fatty acids, the biosynthetic machinery incorporates nearly exclusively cis configuration double bonds. Some food products, such as margarine, may contain trans double bonds because of manipulations during food processing; some evidence suggests that this is a potential health problem, and that food products lacking trans fatty acids are therefore healthier. Monounsaturated fatty acids contain a single double bond, while polyunsaturated fatty acids contain more than one site of unsaturation. The cis bond causes a kink in the fatty acid acyl chain (recall that, unlike single bonds, double bonds do not allow free rotation).
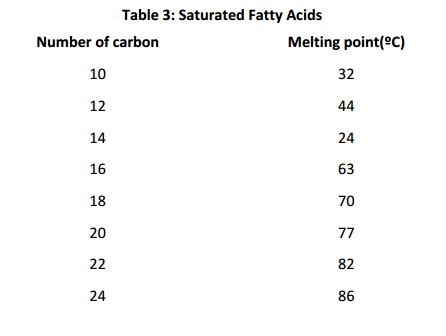
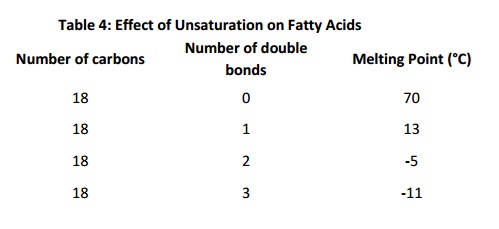
“Partially hydrogenated vegetable oil” is a term often found onfood ingredient labels. It means that some of the fatty acids in the oil were “hydrogenated” (reduced, so that the double bonds were converted to single bonds). Why is this done? Saturated fat and monounsaturated fat forms a solid at room temperature Polyunsaturated fat is (usually) liquid at room temperature. Mixing saturated/monounsaturated/polyunsaturated fatty acids is a way of regulating consistency of food, and also of regulating the consistency of biological systems. This is because, in membranes or in bulk lipid, cis double bonds alter packing density. This decreases van der Waals contacts: the presence of cist double bonds therefore results in lower melting temperature because they result in less regular and less stable structures. The table below shows the effect of chain length and number of cist double bonds on melting temperature (note that the values for melting temperature vary somewhat depending on the reference consulted). Longer chains result in higher melting temperatures;increasing numbers of double bonds decreases melting temperature for fatty acids of a given length
Nomenclature
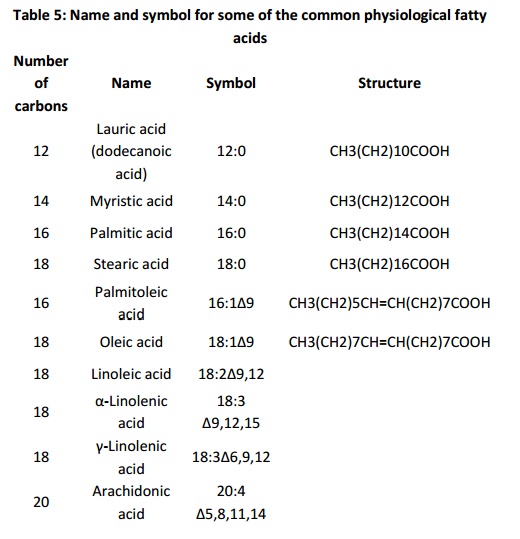
Fatty acids are named using both historical labels and symbols. Both types of nomenclature uniquely define molecules with specific lengths and number and position of double bonds. The table 5 (below)gives the name and symbol for some of the common physiological fatty acids
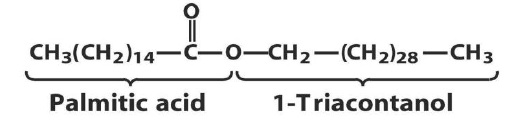
Simple Lipids The second types of simple lipids are waxes:
They are the esters of fatty acids with long chain monohydroxy alcohols 26 to 34 carbons atoms. Waxes are wide-spread in nature and occur usually as mixtures. They form a protective coating on the surfaces of animals and plants. Some insects also secrete waxes. The main constituent of bees wax obtained from the honey comb of bees is myricyl palmitate:
Triacylglycerols:
Triacylglycerol’s (formerly called triglycerides) are complex lipids. Triacylglycerols act as energy storage molecules, especially in adipose tissue; triacylglycerols are also found in lipoproteins. Triacylglycerols are not found in membranes, because they are essentially entirely non-polar. Triacylglycerols consist of three fatty acid molecules forming ester links to glycerol. A triacylglycerol molecule can be comprised of different fatty acids, or of three identical fatty acids. In nature, they are synthesized by enzyme systems, which determine that a centre of asymmetry is created about carbon-2 of the glycerol backbone, so they exist in enantiomeric forms, i.e. with different fatty acids in each position.
A stereo specific numbering system has been recommended to describe these forms. In a Fischer projection of a natural L-glycerol derivative, the secondary hydroxyl group is shown to the left of C-2; the carbon atom above this then becomes C-1 and that below is C-3. The prefix "sn" is placed before the stem name of the compound, when the stereochemistry is defined. Their primary biological function is to serve as a store of energy. As an example, the single molecular species 1,2-dihexadecanoyl-3-(9Z-octadecenoyl)-sn-glycerol is illustrated.
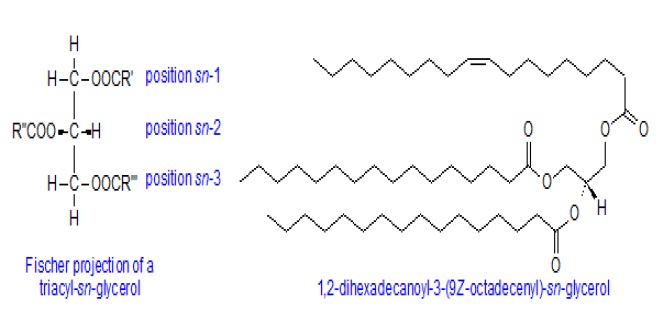
Diacylglycerols (less accurately termed "diglycerides") and monoacylglycerols (monoglycerides) contain two moles and one mole of fatty acids per mole of glycerol, respectively, and exist in various isomeric forms. They are sometimes termed collectively "partial glycerides". Although they are rarely present at greater than trace levels in fresh animal and plant tissues, 1,2-diacyl-sn-glycerols are key intermediates in the biosynthesis of triacylglycerols and other lipids, and they are vital cellular messengers, generated on hydrolysis of phosphatidylinositol and related lipids by a specific phospholipase C. 2-Monoacyl-sn-glycerols are formed as intermediates or end-products of the enzymatic hydrolysis of triacylglycerols; these and other positional isomers are powerful surfactants. 2-Arachidonoylglycerol has important biological properties (as an endocannabinoid).

Acyl migration occurs rapidly in partial glycerides at room temperature, but especially on heating, in alcoholic solvents or in the presence of acid or base, so special procedures are required for their isolation or analysis if the stereochemistry is to be retained. Synthetic 1/3-monoacylglycerols are important in commerce as surfactants.
Phospholipids
There are two classes of phospholipids. The first are the glycerophospholipids, which are themselves subdivided into two groups. The first group, phosphatides, is molecules composed of glycerol substituted with two fatty acid esters (just like in fats) and at the third position a phosphate unit connects to an alcohol.
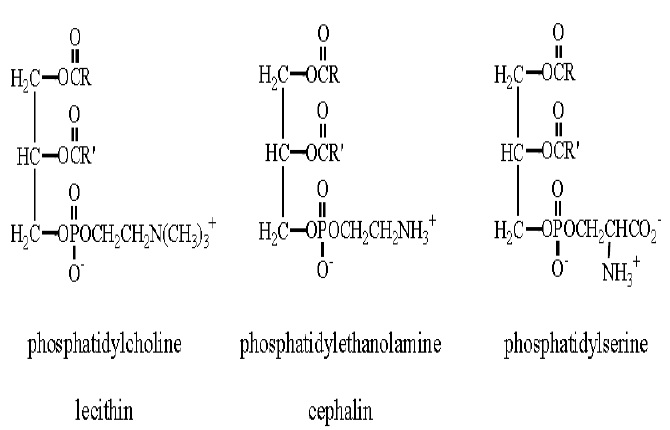
Three alcohols that form phosphatides are choline, ethanolamine, and serine. These compounds are important to the body and are transported as the following phosphatides. The enzymes either cut the molecule free when it is needed or convert it to some other necessary material. The phosphate group and the organic chain attached to it carry electrical charges three phosphatides are components of cell membranes.
Choline is a water-soluble vitamin (as recognized by the Food and Nutrition Board, usually classified as a B vitamin) used to make complex lipids. Phosphatidylcholine is the principal phospholipid of cell membranes. It is also converted to acetyl choline (CH3CO2CH2CH2N (CH3)3+), which is an important neurotransmitter (it carries electrical charges from one nerve cell to another). Choline helps break down homocysteine – a cardiovascular disease risk factor. A lack of this vitamin leads to fatty livers and/or hemorrhagic kidney disease. Serine is the parent of a family of amino acids that also includes glycine and cysteine. Enzymes convert serine (as part of phosphatidylserine) to glycine and cysteine. Serine is also involved in the generation of ethanolamine, which is in turn converted to choline. Interestingly, phosphatidylethanolamine is deficient in Alzheimer’s patients. They also act as a histamine blocker in the body. The other subclass of glycerophospholipids is the plasmalogens. These differ from triacylglycerols by even more than the phosphatides. A generic plasmalogen would look like:
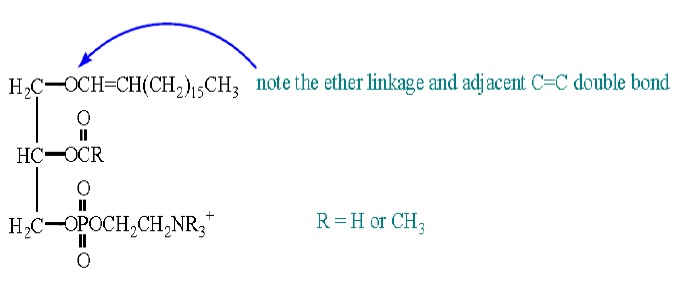
The compound with R = CH3 is called platelet activating factor. It is a strong bronchoconstrictor. It also stimulates other cells to increase their functional and metabolic activities.
The second major class of phospholipids is the sphingolipids. Sphingolipids include the sphingomyelins and cerebrosides. Both are based on the molecule sphingosine. Sphingomyelins have the basic formula:
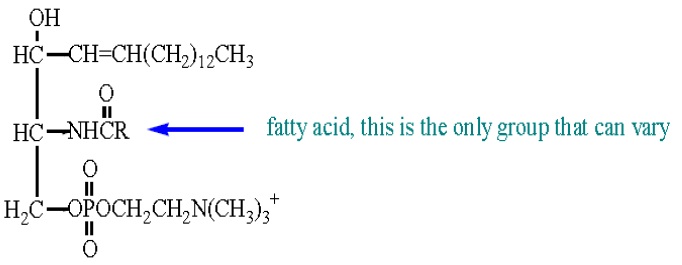
As the name suggests this lipid is affiliated with the myelin sheath surrounding the cells of the central nervous system. Sphingomyelins comprise about 25% of the lipids in the myelin sheath and their role is key to brain function and electrical transmission throughour nervous system. The other type of sphingolipids we are concerned with are cerebrosides, which are not phospholipids. These compounds are again based on attachments to a sphingosine molecule.
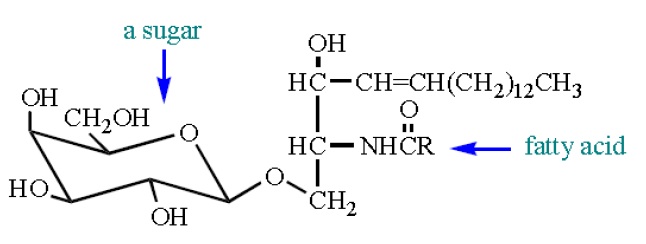
Not surprisingly, these molecules are called glycolipids (cf. glycosides are acetals of sugars). Most of these molecules incorporate Ăź-D-galactose sugars. Cerebrosides are found most commonly in cell membranes in the brain. One cerebroside found outside the brain, a glucocerebroside, is found in the membranes of macrophages (cells that destroy foreign microorganisms). Several disorders are associated with malfunctioning of sphingolipid metabolism. Probably the best known is Tay-Sachs disease, which strikes infants and is typically fatal by age 3. Niemann-Pick disease also strikes infants and is fatal early in life.
Gaucher’s disease and Fabry’s disease strike later in life and are generally less devastating.
Sterols and sterol esters:
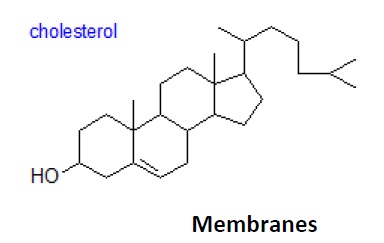
Cholesterol is by far the most common member of a group of steroids in animal tissues; it has a tetra cyclic ring system with a double bond in one of the rings and one free hydroxyl group. It is found both in the Free State, where it has an essential role in maintaining membrane fluidity, and in esterifies form, i.e. as cholesterol esters. Other sterols are present in free and esterifies form in animal tissues, but at trace levelsonly. Cholesterol is the precursor of the bile acids and steroidal hormones. In plants, cholesterol is rarely present in other than small amounts, but such phytosterols as sit sterol, stigma sterol, avenasterol, camp sterol and brassicasterol, and their fatty acid esters are usually found, and they perform a similar function. Hopanoids are related lipids produced by some bacterial species.
Lipids by definition are water insoluble; however, under certain conditions lipids and water are in fact miscible. Consider a typical water-insoluble fatty acid. At elevated pH values, the fatty acid forms soaps with ions such as Na+ and K+. Fatty acids consist of a polar head and a hydrocarbon or no polar tail. Thus, salts of fatty acids are soluble in both polar and no polar solvents. These types of substances are called amphiphatic compounds, i.e., compounds with both polar and nonpolar components. Amphiphatic substances form a number of different structures. Some of these structures are monolayers, micells, and bilayers.
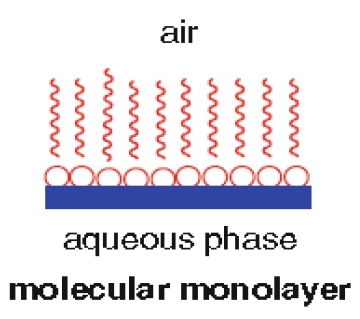
1. A monolayer of lipid may form at the water–lipid interface (Fig. 1.6)
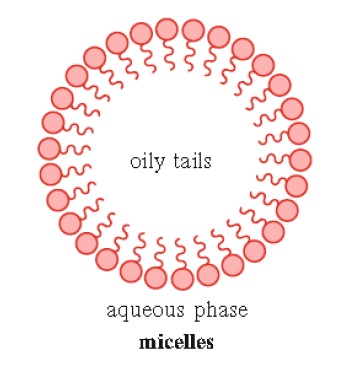
2. Soaps and detergents may form structures known as micelles when they reach a defined concentration in solution. This concentration of amphiphatic compound, known as the critical micelle concentration (CMC), is required before micelle formation can occur. The compounds that form micelles are made up of fatty acids with a single hydrophobic tail (Fig. 1.7)
3. Lipid bilayers may form from lipids that typically contain two hydrophobic tails. The most prominent members of this class are the sphingolipids and glycerophospholipids (Fig. 1.8).
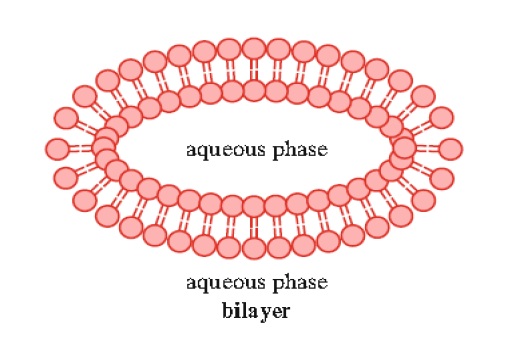
Studies have indicated that the bilayer thickness is approximately 60 A °. Lipid bilayers are structurally very similar to biological membranes and thus their properties have been studied extensively. Experimental bilayers, known as liposomes, have been prepared from phospholipids and sphingolipids by sonication.
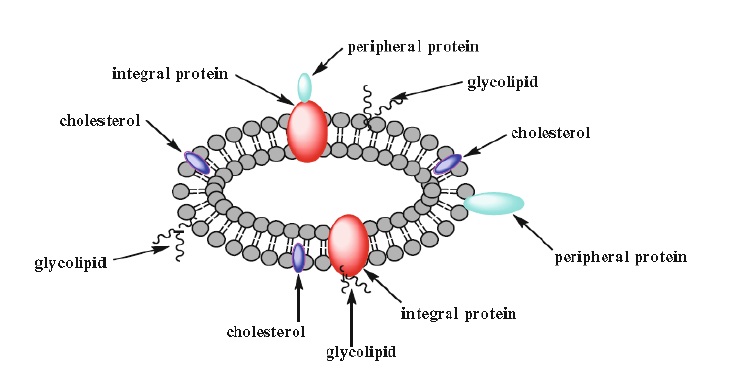
4. Biological membranes will not be discussed in detail; however, simply stated, they are similar to liposomes in that they contain a lipid bilayer, but unlike liposomes they contain two types of proteins. One, the integral proteins, are embedded in the bilayer and the other, the peripheral proteins, are associated either with the surface of the bilayer or with the integral protein itself. Miscellaneous lipids such as cholesterol are also components of biological membranes and affect its fluidity (Fig. 1.9).
Related Topics