Chapter: Medical Physiology: Sports Physiology
Phosphocreatine-Creatine System - Muscles in Exercise
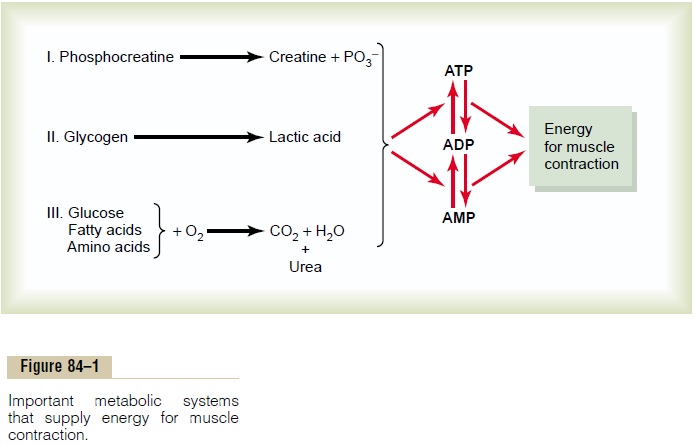
Phosphocreatine-Creatine System
Phosphocreatine (also called creatine phosphate) is another chemical compound that has a high-energy phosphate bond, with the following formula:
Creatine ~ PO3-
This can decompose to creatine and phosphate ion, as shown to the left in Figure 84–1, and in doing so release large amounts of energy. In fact, the high-energy phos-phate bond of phosphocreatine has more energy than the bond of ATP, 10,300 calories per mole in compari-son with 7300. Therefore, phosphocreatine can easily provide enough energy to reconstitute the high-energy bond of ATP. Furthermore, most muscle cells have two to four times as much phosphocreatine as ATP.

A special characteristic of energy transfer from phos-phocreatine to ATP is that it occurs within a small frac-tion of a second. Therefore, all the energy stored in the muscle phosphocreatine is almost instantaneously avail-able for muscle contraction, just as is the energy stored in ATP.
The combined amounts of cell ATP and cell phos-phocreatine are called the phosphagen energy system. These together can provide maximal muscle power for 8 to 10 seconds, almost enough for the 100-meter run.
Thus, the energy from the phosphagen system is used for maximal short bursts of muscle power.
Glycogen-Lactic Acid System.The stored glycogen inmuscle can be split into glucose and the glucose then used for energy. The initial stage of this process, called glycolysis, occurs without use of oxygen and, therefore,is said to be anaerobic metabolism. During glycolysis, each glucose molecule is split into two pyruvic acid molecules, and energy is released to formfour ATP molecules for each original glucose molecule. Ordinarily, the pyruvic acid then enters the mitochondria of the muscle cells and reacts with oxygen to form still many more ATP molecules. However, when there is insufficient oxygen for this second stage (the oxidative stage) of glucose metabolism to occur, most of the pyruvic acid then is converted into lactic acid, which diffuses out of the muscle cells into the interstitial fluid and blood. Therefore, much of the muscle glycogen is transformed to lactic acid, but in doing so, considerable amounts of ATP are formed entirely without the consumption of oxygen.
Another characteristic of the glycogen-lactic acid system is that it can form ATP molecules about 2.5 times as rapidly as can the oxidative mechanism of the mitochondria. Therefore, when large amounts of ATP are required for short to moderate periods of muscle contraction, this anaerobic glycolysis mechanism can be used as a rapid source of energy. It is, however, only about one half as rapid as the phosphagen system. Under optimal conditions, the glycogen-lactic acid system can provide 1.3 to 1.6 minutes of maximal muscle activity in addition to the 8 to 10 seconds pro-vided by the phosphagen system, although at somewhat reduced muscle power.
Aerobic System. The aerobic system is the oxidation offoodstuffs in the mitochondria to provide energy. That is, as shown to the left in Figure 84–1, glucose, fatty acids, and amino acids from the foodstuffs—after some inter-mediate processing—combine with oxygen to release tremendous amounts of energy that are used to convert AMP and ADP into ATP.
In comparing this aerobic mechanism of energy supply with the glycogen-lactic acid system and the phosphagen system, the relativemaximal rates of powergeneration in terms of moles of ATP generation perminute are the following:

When comparing the same systems for endurance, the relative values are the following:

Thus, one can readily see that the phosphagen system is the one used by the muscle for power surges of a few seconds, and the aerobic system is required for pro-longed athletic activity. In between is the glycogen-lactic acid system, which is especially important for giving extra power during such intermediate races as the 200-to 800-meter runs.
What Types of Sports Use Which Energy Systems? By consid-ering the vigor of a sports activity and its duration, one can estimate closely which of the energy systems is used for each activity. Various approximations are presented in Table 84–1.
Recovery of the Muscle Metabolic Systems After Exercise. Inthe same way that the energy from phosphocreatine can be used to reconstitute ATP, energy from the glycogen-lactic acid system can be used to reconstitute both phos-phocreatine and ATP. And then energy from the oxidative metabolism of the aerobic system can be used to reconstitute all the other systems-the ATP, the phos-phocreatine, and the glycogen-lactic acid system.
Reconstitution of the lactic acid system means mainly the removal of the excess lactic acid that has accumu-lated in all the fluids of the body. This is especially important because lactic acid causes extreme fatigue.
When adequate amounts of energy are available from oxidative metabolism, removal of lactic acid is achieved in two ways: (1) A small portion of it is converted back into pyruvic acid and then metabolized oxidatively by all the body tissues. (2) The remaining lactic acid is reconverted into glucose mainly in the liver, and the glucose in turn is used to replenish the glycogen stores of the muscles.
Recovery of the Aerobic System After Exercise. Even duringthe early stages of heavy exercise, a portion of one’s aerobic energy capability is depleted. This results from two effects: (1) the so-called oxygen debt and (2) deple-tion of the glycogen stores of the muscles.
Oxygen Debt. The body normally contains about 2 litersof stored oxygen that can be used for aerobic metabo-lism even without breathing any new oxygen. This stored oxygen consists of the following: (1) 0.5 liter in the air of the lungs, (2) 0.25 liter dissolved in the body fluids, (3) 1 liter combined with the hemoglobin of the blood, and (4) 0.3 liter stored in the muscle fibers them-selves, combined mainly with myoglobin, an oxygen-binding chemical similar to hemoglobin.
In heavy exercise, almost all this stored oxygen is used within a minute or so for aerobic metabolism. Then, after the exercise is over, this stored oxygen must be replenished by breathing extra amounts of oxygen over and above the normal requirements. In addition, about 9 liters more oxygen must be consumed to provide for reconstituting both the phosphagen system and the lactic acid system. All this extra oxygen that must be “repaid,” about 11.5 liters, is called the oxygen debt.
Figure 84–2 shows this principle of oxygen debt. During the first 4 minutes of the figure, the person exer-cises heavily, and the rate of oxygen uptake increases
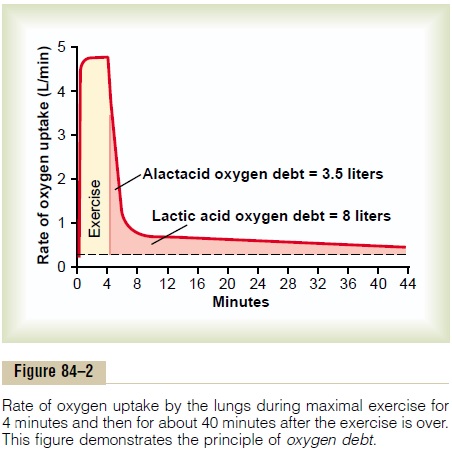
more than 15-fold. Then, even after the exercise is over, the oxygen uptake still remains above normal, at first very high while the body is reconstituting the phospha-gen system and repaying the stored oxygen portion of the oxygen debt, and then for another 40 minutes at a lower level while the lactic acid is removed. The early portion of the oxygen debt is called the alactacid oxygendebt and amounts to about 3.5 liters. The latter portionis called the lactic acid oxygen debt and amounts to about 8 liters.
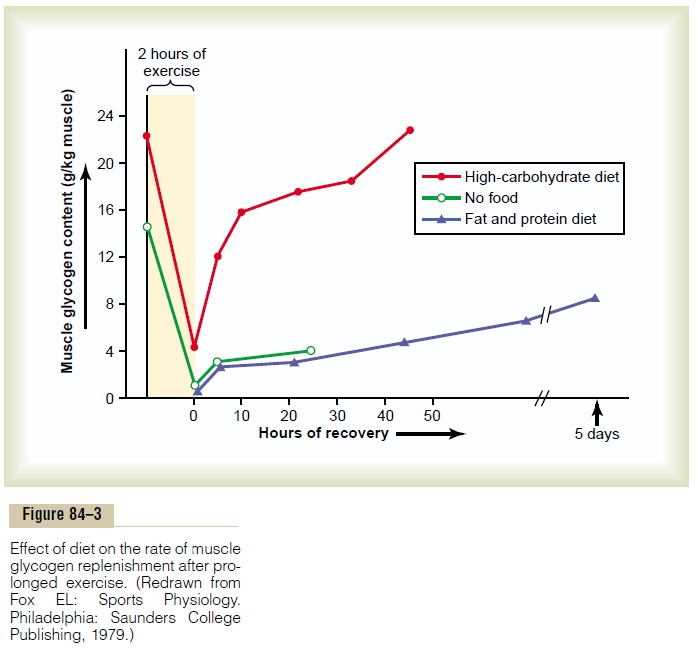
Recovery of Muscle Glycogen. Recovery from exhaustivemuscle glycogen depletion is not a simple matter. This often requires days, rather than the seconds, minutes, or hours required for recovery of the phosphagen and lactic acid metabolic systems. Figure 84–3 shows this recovery process under three conditions: first, in people on a high-carbohydrate diet; second, in people on a high-fat, high-protein diet; and third, in people with no food. Note that on a high-carbohydrate diet, full recov-ery occurs in about 2 days. Conversely, people on a high-fat, high-protein diet or on no food at all show very little recovery even after as long as 5 days. The messages of this comparison are (1) that it is important for an athlete to have a high-carbohydrate diet before a grueling ath-letic event and (2) not to participate in exhaustive exer-cise during the 48 hours preceding the event.
Related Topics