Chapter: Basic & Clinical Pharmacology : Histamine, Serotonin, & the Ergot Alkaloids
Basic Pharmacology of Histamine
BASIC PHARMACOLOGY OF HISTAMINE
Chemistry & Pharmacokinetics
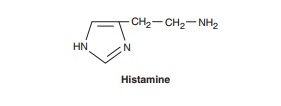
Histamine
occurs in plants as well as in animal tissues and is a component of some venoms
and stinging secretions.
Histamine
is formed by decarboxylation of the amino acid L-histidine, a reaction catalyzed
in mammalian tissues by the enzyme histidine decarboxylase. Once formed,
histamine is either stored or rapidly inactivated. Very little histamine is
excreted unchanged. The major metabolic pathways involve conversion to N-methylhistamine, methylimidazoleacetic
acid, and imida-zoleacetic acid (IAA). Certain neoplasms (systemic
mastocytosis, urticaria pigmentosa, gastric carcinoid, and occasionally
myelog-enous leukemia) are associated with increased numbers of mast cells or
basophils and with increased excretion of histamine and its metabolites.
Most
tissue histamine is sequestered and bound in granules (vesicles) in mast cells
or basophils; the histamine content of many tissues is directly related to
their mast cell content. The bound form of histamine is biologically inactive,
but as noted below, many stimuli can trigger the release of mast cell
histamine, allow-ing the free amine to exert its actions on surrounding
tissues. Mast cells are especially rich at sites of potential tissue
injury—nose, mouth, and feet; internal body surfaces; and blood vessels,
par-ticularly at pressure points and bifurcations.Non-mast cell histamine is
found in several tissues, including the brain, where it functions as a
neurotransmitter. Strong evi-dence implicates endogenous neurotransmitter
histamine in many brain functions such as neuroendocrine control,
cardiovascular regulation, thermal and body weight regulation, and sleep and
arousal.
A
second important nonneuronal site of histamine storage and release is the
enterochromaffin-like (ECL) cells of the fundus of the stomach. ECL cells
release histamine, one of the primary gas-tric acid secretagogues, to activate
the acid-producing parietal cells of the mucosa .
Storage & Release of Histamine
The
stores of histamine in mast cells can be released through sev-eral mechanisms.
A. Immunologic Release
Immunologic
processes account for the most important pathophys-iologic mechanism of mast
cell and basophil histamine release. These cells, if sensitized by IgE
antibodies attached to their surface membranes, degranulate explosively when
exposed to the appro-priate antigen (see Figure 55–5, effector phase). This
type of release also requires energy and calcium. Degranulation leads to the
simultaneous release of histamine, adenosine triphosphate (ATP), and other
mediators that are stored together in the gran-ules. Histamine released by this
mechanism is a mediator in immediate (type I) allergic reactions, such as hay
fever and acute urticaria. Substances released during IgG- or IgM-mediated
immune reactions that activate the complement cascade also release histamine
from mast cells and basophils.
By
a negative feedback control mechanism mediated by H2 receptors,
histamine appears to modulate its own release and that of other mediators from
sensitized mast cells in some tissues. In humans, mast cells in skin and
basophils show this negative feed-back mechanism; lung mast cells do not. Thus,
histamine may act to limit the intensity of the allergic reaction in the skin
and blood.
Endogenous
histamine has a modulating role in a variety of inflammatory and immune responses.
Upon injury to a tissue, released histamine causes local vasodilation and
leakage of plasma-containing mediators of acute inflammation (complement,
C-reactive protein) and antibodies. Histamine has an active chemotactic
attraction for inflammatory cells (neutrophils, eosino-phils, basophils,
monocytes, and lymphocytes). Histamine inhib-its the release of lysosome
contents and several T- and B-lymphocyte functions. Most of these actions are
mediated by H2 or H4 recep-tors. Release of peptides from
nerves in response to inflammation is also probably modulated by histamine, in
this case acting through presynaptic H3 receptors.
B. Chemical and Mechanical Release
Certain
amines, including drugs such as morphine and tubocura-rine, can displace
histamine from its bound form within cells. This type of release does not
require energy and is not associated with mast cell injury or degranulation.
Loss of granules from the mast cell also releases histamine, since sodium ions
in the extracellular fluid rapidly displace the amine from the complex.
Chemical and mechanical mast cell injury causes degranulation and histamine
release. Compound 48/80, an
experimental drug, selectively releases histamine from tissue mast cells by an
exocytotic degranu-lation process requiring energy and calcium.
Pharmacodynamics
A. Mechanism of Action
Histamine
exerts its biologic actions by combining with specific cellular receptors
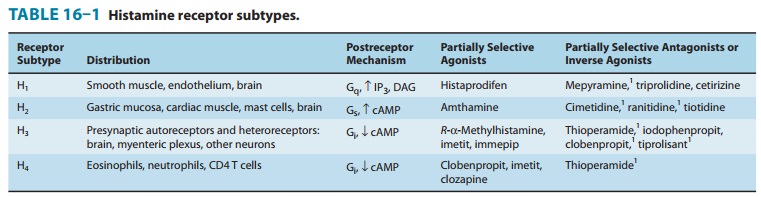
located on the surface membrane. Four different histamine receptors have been
characterized and are designated H1–H4; they are
described in Table 16–1. Unlike the other amine transmitter receptors discussed
previously, no subfamilies have been found within these major types, although
different splice variants of several receptor types have been described.
All
four receptor types have been cloned and belong to the large superfamily of
receptors having seven membrane-spanning regions and coupled with G proteins
(GPCR). The structures of the H1 and H2 receptors differ
significantly and appear to be more closely related to muscarinic and 5-HT1
receptors, respectively, than to each other. The H4 receptor has
about 40% homology with the H3 receptor but does not seem to be
closely related to any other histamine receptor. All four histamine receptors
have been shown to have constitutive activity in some systems; thus, some
antihistamines previously con-sidered to be traditional pharmacologic
antagonists must now be considered to be inverse agonists. Indeed, many first-
and second-generation H1 blockers
function as inverse agonists. Furthermore, a single molecule may be an
agonist at one histamine receptor and an antagonist or inverse agonist at
another. For example, clobenpropit, an agonist at H4 receptors, is
an antagonist or inverse agonist at H3 receptors (Table 16–1).
In
the brain, H1 and H2 receptors are located on
postsynaptic membranes, whereas H3 receptors are predominantly
presynaptic. Activation of H1 receptors, which are present in
endothelium, smooth muscle cells, and nerve endings, usually elicits an
increase in phosphoinositol hydrolysis and an increase in inositol
trisphosphate (IP3) and intracellular calcium. Activation of H2
receptors, present in gastric mucosa, cardiac muscle cells, and some immune
cells, increases intracellular cyclic adenosine monophosphate (cAMP) via Gs.
Like the β2 adrenoceptor, under certain circumstances the H2
receptor may couple to Gq, activating the IP3-DAG
(inositol 1,4,5-trisphosphate-diacylglycerol) cascade. Activation of H3
receptors decreases transmitter release from histaminergic and other neurons,
probably mediated by a decrease in calcium influx through N-type calcium
channels in nerve endings. H4 receptors are found mainly on
leukocytes in the bone marrow and circulating blood. H4 recep-tors
appear to have very important chemotactic effects on eosino-phils and mast
cells. In this role, they seem to play a part in inflammation and allergy. They
may also modulate production of these cell types and they may mediate, in part,
the previously recog-nized effects of histamine on cytokine production.
B. Tissue and Organ System Effects of Histamine
Histamine
exerts powerful effects on smooth and cardiac muscle, on certain endothelial
and nerve cells, on the secretory cells of the stomach, and on inflammatory
cells. However, sensitivity to hista-mine varies greatly among species. Guinea
pigs are exquisitely sensitive; humans, dogs, and cats somewhat less so; and
mice and rats very much less so.
1. Nervous system—Histamine is a
powerful stimulant of sensory nerve endings, especially those mediating pain
and itching. This H1-mediated effect is an important component of
the urti-carial response and reactions to insect and nettle stings. Some
evidence suggests that local high concentrations can also depolar-ize efferent
(axonal) nerve endings (see Triple Response, item 8 in this list). In the
mouse, and probably in humans, respiratory neu-rons signaling inspiration and
expiration are modulated by H1 receptors. H1 and H3
receptors play important roles in appetite and satiety; antipsychotic drugs
that block these receptors cause significant weight gain . Presynaptic H3
receptors play important roles in modulating release of several transmitters in
the nervous system. H3 agonists reduce the release of
acetylcho-line, amine, and peptide transmitters in various areas of the brain
and in peripheral nerves.
2. Cardiovascular system—In humans, injection or infusionof histamine causes a decrease in systolic and diastolic blood pres-sure and an increase in heart rate. The blood pressure changes are caused by the direct vasodilator action of histamine on arterioles and precapillary sphincters; the increase in heart rate involves both stimulatory actions of histamine on the heart and a reflex tachy-cardia. Flushing, a sense of warmth, and headache may also occur during histamine administration, consistent with the vasodilation.
Vasodilation
elicited by small doses of histamine is caused by H1-receptor
activation and is mediated mainly by release of nitric oxide from the
endothelium . The decrease in blood pressure is usually accompanied by a reflex
tachycardia. Higher doses of histamine activate the H2-mediated cAMP
pro-cess of vasodilation and direct cardiac stimulation. In humans, the cardiovascular
effects of small doses of histamine can usually be antagonized by H1-receptor
antagonists alone.
Histamine-induced edema results from the action of the amine on H1 receptors in the vessels of the microcirculation, especially the postcapillary vessels. The effect is associated with the separation of the endothelial cells, which permits the transudation of fluid and molecules as large as small proteins into the perivascular tissue. This effect is responsible for urticaria (hives), which signals the release of histamine in the skin. Studies of endothelial cells suggest that actin and myosin within these cells cause contraction, resulting in separa-tion of the endothelial cells and increased permeability.
Direct
cardiac effects of histamine include both increased con-tractility and
increased pacemaker rate. These effects are mediated chiefly by H2
receptors. In human atrial muscle, histamine can also decrease contractility;
this effect is mediated by H1 receptors. The physiologic
significance of these cardiac actions is not clear. Some of the cardiovascular
signs and symptoms of anaphylaxis are due to released histamine, although
several other mediators are involved and appear to be more important than
histamine in humans.
3. Bronchiolar smooth muscle—In both humans and
guineapigs, histamine causes bronchoconstriction mediated by H1
recep-tors. In the guinea pig, this effect is the cause of death from
his-tamine toxicity, but in humans with normal airways, broncho-constriction
following small doses of histamine is not marked. However, patients with asthma
are very sensitive to histamine. The bronchoconstriction induced in these
patients probably represents a hyperactive neural response, since such patients
also respond excessively to many other stimuli, and the response to
histaminecan be blocked by autonomic blocking drugs such as ganglion blocking
agents as well as by H1-receptor antagonists . Although methacholine
provocation is more commonly used, tests using small doses of inhaled histamine
have been used in the diagnosis of bronchial hyperreactivity in patients with
suspected asthma or cystic fibrosis. Such individuals may be 100 to 1000 times
more sensitive to histamine (and methacholine) than are normal subjects.
Curiously, a few species (eg, rabbit) respond tohistamine with bronchodilation, reflecting the dominance of
the H2 receptor in their airways.
4. Gastrointestinal tract smooth
muscle—Histamine
causescontraction of intestinal smooth muscle, and histamine-induced
contraction of guinea pig ileum is a standard bioassay for this amine. The
human gut is not as sensitive as that of the guinea pig, but large doses of histamine may
cause diarrhea, partly as a result of this effect. This action of histamine is
mediated by H1 receptors.
5. Other smooth muscle organs—In humans, histamine
gen-erally has insignificant effects on the smooth muscle of the eye
andgenitourinary tract. However, pregnant women suffering anaphylactic
reactions may abort as a result of histamine-induced contractions, and in some
species the sensitivity of the uterus is sufficient to form the basis for a
bioassay.
6. Secretory tissue—Histamine has long
been recognized as apowerful stimulant of gastric acid secretion and, to a
lesser extent, of gastric pepsin and intrinsic factor
production. The effect is caused by activation of H2 receptors on
gastric parietal cells and is associated with increased adenylyl cyclase
activity, cAMP concen-tration, and intracellular Ca2+ concentration. Other
stimulants of gastric acid secretion such as acetylcholine and gastrin do not
increase cAMP even though their maximal effects on acid output can be
reduced—but not abolished—by H2-receptor antagonists.
Histamine
also stimulates secretion in the small and large intes-tine. In contrast, H3-selective
histamine agonists inhibit acid
secre-tion stimulated by food or pentagastrin in several species.
Histamine
has much smaller effects on the activity of other glandular tissue at ordinary
concentrations. Very high concentra-tions can cause adrenal medullary
discharge.
7. Metabolic effects—Recent studies of H3-receptor
knockoutmice demonstrate that absence of this receptor results in animals with
increased food intake, decreased energy expenditure, and obesity. They also
show insulin resistance and increased blood levels of leptin and insulin. It is
not yet known whether the H3 receptor has a similar role in humans,
but intensive research is underway to determine whether H3 agonists
are useful in the treatment of obesity.
8. The “triple response”—Intradermal injection
of histaminecauses a characteristic red spot, edema, and flare response that
was first described many years ago. The effect involves three separate cell
types: smooth muscle in the microcirculation, capillary or venular endothelium,
and sensory nerve endings. At the site of injection, a reddening appears owing
to dilation of small vessels, followed soon by an edematous wheal at the
injection site and a red irregular flare surrounding the wheal. The flare is
said to be caused by an axon reflex. A sensation of itching may accompany these
effects.
Similar
local effects may be produced by injecting histamine liberators (compound
48/80, morphine, etc) intradermally or by applying the appropriate antigens to
the skin of a sensitized per-son. Although most of these local effects can be
prevented by adequate doses of an H1-receptor–blocking agent, H2
and H3 receptors may also be involved.
9. Other effects possibly
mediated by histamine receptors—In addition to the local stimulation of
peripheralpain nerve endings via H3 and H1 receptors,
histamine may play a role in nociception in the central nervous system. Burimamide, an early candidate for H2-blocking
action, and newer analogs with no effect on H1, H2, or H3
receptors, have been shown to have significant analgesic action in rodents when
administered into the central nervous system. The analgesia is said to be
comparable to that produced by opioids, but tolerance, respiratory depression,
and constipation have not been reported. Although the mecha-nism of this action
is not known, these compounds may represent an important new class of
analgesics.
Other Histamine Agonists
Small
substitutions on the imidazole ring of histamine signifi-cantly modify the
selectivity of the compounds for the histamine receptor subtypes. Some of these
are listed in Table 16–1.
Related Topics